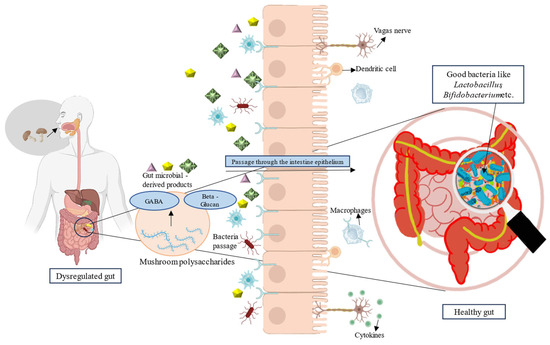
ハイライト
- この総説は、虚血性脳卒中におけるピロトーシスの最新の分子機構を要約したものである。
- このレビューは、虚血性脳卒中におけるピロトーシスの調節因子としての天然植物成分の役割を要約しています。
- 虚血性脳卒中におけるピロトーシスの制御は、潜在的な治療戦略として提案されています。
抽象的な
虚血性脳卒中 (IS) は、死亡および障害の主な原因の 1 つです。その病因は完全には明らかではなく、炎症カスケードはその主な原因の 1 つです。 病理学的プロセス。 IS の現在の臨床実践は、 血液供給 IS後はできるだけ早く虚血領域に到達します。 血栓溶解療法 ニューロンの活力と機能を保護します。しかし、血液再灌流は虚血損傷をさらに加速し、虚血再灌流損傷を引き起こします。脳虚血再灌流障害の病理学的プロセスには複数のメカニズムが関与しており、正確なメカニズムはまだ完全には解明されていません。 パイロトーシス、新たに発見された炎症性の形態 プログラムされた細胞死、炎症の開始と進行に重要な役割を果たします。これは、炎症促進性のプログラムされた死であり、 カスパーゼ カスパーゼ 1/4/5/11 は細胞の膨張と破裂を引き起こし、炎症因子 IL-1β と IL-18 を放出し、炎症カスケードを誘導します。最近の研究では、パイロトーシスとその媒介炎症反応が虚血性脳損傷を悪化させる重要な因子であり、パイロトーシスを抑制することで虚血性脳損傷が軽減される可能性があることが示されています。さらに、研究により、天然の植物成分がパイロトーシスを制御する効果がある可能性があることが判明しました。したがって、この総説は、パイロトーシスの分子機構と虚血性脳卒中におけるその役割を要約するだけでなく、パイロトーシスに関する参考情報を提供するために、パイロトーシスの調節因子としての天然植物成分の役割も要約する。 処理 将来のISのこと。
キーワード
1. はじめに
世界人口の高齢化に伴い、 脳血管疾患 脳卒中などの感染者数は世界第2位となり、人類への脅威は増大している [1]。脳卒中には以下が含まれます 虚血性脳卒中 (IS)と 出血性脳卒中、最大 87% の脳卒中は虚血性です [2]。 IS は一般的な臨床緊急事態であり、その発生率は年々増加しています。 IS の現在の臨床実践は、 血液供給 IS後はできるだけ早く虚血領域に到達します。 血栓溶解療法 ニューロンの活力と機能を保護する [3], [4]。しかし、血流回復後の虚血再灌流障害(IRI)は、多くの場合、より深刻な損傷を引き起こします。 脳組織 さらに、患者の予後不良や機能不全を引き起こす重要な要因となります。 [5]。この世界的疫学調査は、世界204の国と地域における1990年から2019年までのデータを分析した。彼らの報告書によると、脳卒中は依然として死因の第 2 位(総死亡数の 11.6%)であり、死亡および障害の原因の第 3 位である(総死亡数の 5.7%)。 障害調整後の生存年数 (DALYs)]に次ぐ2位です。 新生児疾患 (7.3% DALY) および 虚血性心疾患 (7.2% DALY)、脳卒中患者の数は依然として増加しています [6], [7]、特に中国では。中国は世界で最も脳卒中リスクが高い国となっており、住民の脳卒中リスクは39.3%に達し、中国人男性の脳卒中リスクも世界で最も高く、41%を超えている。 ISは中国人住民の死因の第3位(1990年時点)から第1位(2017年時点)に上昇した [8]。現在の研究では、脳 IRI (CIRI) の病理学的メカニズムは主に次のとおりであることが示唆されています。 酸化ストレス けが、 炎症性損傷、ミトコンドリア損傷、オートファジー、 アポトーシス [9], [10], [11], [12], [13], [14]。しかし、これらは病理学的メカニズムを説明するにはまだ不十分です。 シリ。例えば、CIRI後の神経細胞死は主にアポトーシスによって引き起こされると考えられており、神経細胞保護の方向性は主に抗アポトーシスである。ただし、アポトーシスは炎症とは直接関係ありません [15], [16]しかし、CIRIにはかなりの炎症反応が存在します。 [16]。したがって、ISの病態生理学的メカニズムをさらに解明し、CIRIの病因をさらに調査する必要があります。
パイロトーシス、 としても知られている 細胞炎症性 壊死は、近年新たに発見され確認されたプログラム細胞死です。 [17]。パイロトーシスは炎症に依存しています カスパーゼ (主にカスパーゼ-1、4、5、11)そして多数の炎症促進因子の放出を伴います。それは、細胞が成長するまで細胞が継続的に増殖することによって現れます。 膜の破裂、細胞内容物の放出と強い炎症反応の活性化を引き起こします。 [17], [18]。の 形態的特徴、パイロトーシスの発生と制御メカニズムは、アポトーシスやネクローシスなどの他の細胞死方法とは異なります [18]。パイロトーシスは、感染症、神経系関連疾患、アテローム性動脈硬化症、悪性腫瘍などのさまざまな疾患の発生と発症に広く関与しています。 [19], [20], [21]。最新の研究では、ピロトーシスとそれを介した炎症反応が症状に関与していることが示されています。 病理学的プロセス パイロトーシスの活性化を防ぐことは、炎症カスケードを抑制し、虚血性脳損傷を軽減するのに有益です。 [22], [23]。これに基づいて、パイロトーシスの研究は、 処理 ISの。パイロトーシスを制御すると、ISの致死率が低下し、ニューロンの生存率が向上し、IS患者の症状が改善される可能性があります。現在、 薬物 力を発揮する 神経保護的な パイロトーシス経路を調節することによる効果が開発されています。特に、IS における血管ニューロン単位のパイロトーシスを制御する天然植物成分に関する研究が数多く行われています。私たちの以前の研究でも、 サポニン [24], [25], [26] および多成分化合物 [27], [28] を規制できる 生物学的プロセス ISにおけるパイロトーシスの症状。したがって、この総説は、パイロトーシスの分子機構と虚血性脳卒中におけるその役割を要約するだけでなく、将来のパイロトーシス関連薬剤開発のためのリード化合物または天然植物成分を提供するために、パイロトーシスの調節因子としての天然植物成分の役割も要約する。
2. パイロトーシス
2.1.パイロトーシスの特徴
パイロトーシス の形式です プログラムされた細胞死 炎症誘発物質に依存する カスパーゼ、膜貫通孔の形成、細胞膜の膨張と破裂、炎症誘発性内容物の放出を特徴とします。 [29]。アポトーシスの過程では、細胞は核の断片化と細胞膜の完全性を持ち、周囲の組織に炎症反応を引き起こしませんが、パイロトーシスの過程では細胞は完全な核の形態と細胞を持ちます。 膜の破裂、末梢炎症を引き起こす [30]。ピロトーシスが起こると、細胞膜が徐々に破れて1~2nmの細胞膜孔が形成され、細胞内炎症因子やその他の物質などの細胞内容物が細胞膜の外に放出されます。膜透過性色素 臭化エチジウム 細胞がパイロトーシスを起こすかどうかを観察するために使用できます [31]。さらに、壊死の発生はプロテインキナーゼ様の混合系列に依存します。 ドメインタンパク質 (MLKL)、細胞に入るイオンに対して選択的であり、核クロマチンは凝集性または端に凝集しています。パイロトーシスは、核分裂を引き起こす非選択的タンパク質である孔形成タンパク質 GSDMD に依存しています。 濃縮症 そして DNAの断片化 [32]。ピロトーシス、アポトーシス、ネクロトーシス、およびネクロトーシスには類似点と相違点があります。 フェロトーシス原因や細胞の変化など。詳細は、 表1.
表1。パイロトーシス、アポトーシス、ネクロトーシスの類似点と相違点。
| 特徴 | 署名コンポーネント | 細胞の変化 | 一般的な原因 | 阻害剤 | 参照 |
|---|---|---|---|---|---|
| パイロトーシス | ピロプトソーム、インフラマソーム | 孔の形成、細胞の膨張、原形質膜の破裂、クロマチンの凝縮、DNA の断片化と核の完全性など。 | DAMP、PAMP、感染症 | GSDMD阻害剤 | [19], [33], [34], [35], [36] |
| アポトーシス | アポトソーム | 細胞は無傷の膜、細胞膜の水疱形成、クロマチン凝縮、DNA 断片化などを伴って縮小します。 | 生理学的状態の遺伝子制御 | カスパーゼファミリー、Bcl-2ファミリー、p53などを制御 | [37], [38], [39] |
| ネクロプトーシス | ネクロソーム | 細胞の膨張、細胞膜の破裂、細胞小器官の膨張、クロマチンの凝縮など。 | 重傷 | Nec-1、NSAなど | [33], [40], [41], [42], [43] |
| フェロトーシス | NRF2、GPX4、ACSL4など | 主にミトコンドリアで発生し、ミトコンドリアのクリステが減少し、膜が凝縮し、外膜が破壊されます。 | Fe2+過負荷とROS | GSH、デフェロキサミン、リプロスタチン-1、フェロスタチン-1など | [44], [45], [46], [47] |
2.2.パイロトーシスの分子機構
の形成 インフラマソーム 損傷後のパイロトーシスの重要な物質です。損傷を受けると、体は病原体関連分子パターン (PAMP) および損傷関連分子パターン (DAMP) に関連するパターン信号を活性化することがあります。 [48]。信号刺激が起こると、細胞内 プロモータータンパク質 ヌクレオチド結合オリゴマー化ドメイン様受容体 pyrin ドメイン 3 (NLRP3) を含むものは、オリゴマー化を通じてインフラマソーム アダプター分子 ASC を使用して大量のプロカスパーゼ 1 を動員できます。 [49]、インフラマソーム凝集タンパク質の形成。プロカスパーゼ-1 はカスパーゼ-1 の前駆体であり、通常、プロカスパーゼ-1 は次のような形で体内に存在します。 チモーゲン。刺激シグナルがプロカスパーゼ 1 に伝達されると、自己加水分解によって P20 および P10 サブユニットが生成されます。このサブユニットは最初に ヘテロ二量体 それは機能できず、集合して 四量体 カスパーゼ-1の活性を促進し、カスパーゼ-1の活性化をもたらします。 [50]。カスパーゼ-1 が活性化されると、活性型カスパーゼ-1 は細胞膜孔の形成と細胞膜の急速な溶解を引き起こし、炎症反応を形成します。一方、カスパーゼ-1は、IL-1βおよびIL-18の前駆体、プロIL-1βおよびプロIL-18を誘導して、成熟プロセスを促進することもできます。成熟したIL-1βとIL-18は細胞外に放出され、それによってより多くのインフラマソームを動員します。インフラマソームが凝集すると炎症反応はさらに悪化し、組織損傷が悪化します。 IL-1βが細胞から放出されると、リンパの流れに乗って炎症因子が隣接組織に広がり、炎症反応がさらに悪化します。大量の IL-18 の分泌は Th1 および Th2 免疫応答を引き起こし、免疫応答を刺激して役割を果たします [51].
2.2.1.正規のパイロトーシス経路
カスパーゼ-1 は標準的なパイロトーシス経路における重要なタンパク質であり、インフラマソームはカスパーゼ-1 の活性化において重要な役割を果たします。 [52]。インフラマソームは自然免疫系の重要な部分であり、センサータンパク質、ASC、およびプロカスパーゼ-1 で構成される複数タンパク質複合体です。刺激を受けた細胞質に存在します。 免疫細胞 細胞外刺激信号を感知できる [53], [54]。インフラマソームは、ヌクレオチド結合オリゴマー化ドメイン様受容体 (NLR) を含むインフラマソームに分類されます。 黒色腫 2 (AIM2) インフラマソームおよび NLRC4 インフラマソーム [55], [56]。プリン作動性 2 × 7 (P2X7) 受容体は、ATP 依存性膜貫通イオンチャネル受容体であり、 ミクログリア インフラマソーム活性化の重要な要素です。研究では、細胞外ATPがK+の排出を活性化することで調節できることが示されています。 P2X7受容体、NLR ファミリー、ピリンドメイン含有 3 (NLRP3) の活性化を誘導します。 [57]。 NLRP3 インフラマソームは現在最も研究されています。 NLRP3 インフラマソームの活性化には 2 段階の反応が必要です。反応の開始、微生物または内因性因子の核因子-κB (NF-κB) の核内への流入、および NLRP3 およびプロ IL-1β 発現の上方制御です。 [58], [59]。免疫細胞が DAMP によって刺激されると、センサータンパク質が ASC を介してプロカスパーゼ 1 を補充し、その後、自己加水分解によって活性化されます。成熟カスパーゼ-1 は、下流で不活性なプロ IL-1β およびプロ IL-18 を活性型 IL-1β および IL-18 にプロセスします。 [60]。一方、Caspase-1 は、細孔形成活性により下流の GSDMD タンパク質を GSDMD-N フラグメントと GSDMD-C フラグメントに切断します。 GSDMD-N フラグメントは、特異的に認識して結合します。 膜脂質 細胞膜の内側で細胞内および細胞外の浸透圧の変化を引き起こし、その結果、細胞の膨張と破裂、炎症性因子IL-1βおよびIL-18の放出が引き起こされ、炎症反応と細胞ピロトーシスを誘発します。 [61].
2.2.2.非標準的なパイロトーシス経路
カスパーゼファミリーは、異なる機能に応じて主にアポトーシス関連と炎症関連の 2 つのカテゴリーに分類されます。カスパーゼ-3/2/10 は主にアポトーシスを媒介し、カスパーゼ-1/4/5/11 は炎症と自然免疫応答の重要なメディエーターです。 [62], [63]。非標準的なパイロトーシス経路はカスパーゼ-4/5/11の活性化に依存しますが、インフラマソームはIL-1βとIL-18の成熟に必須ではありません [64]。パイロトーシスの非古典的経路は以下によって誘発されます。 リポ多糖類 (LPS)、グラム陰性菌の細胞壁成分。 LPS はヒト カスパーゼ 4/5 およびマウス カスパーゼ 11 に直接結合できます。これらのカスパーゼは、LPS のセンサータンパク質とエフェクター分子の両方として機能します。活性化されたカスパーゼ-4/5/11 は GSDMD を直接切断し、パイロトーシスを誘発します。 [65]。さらに、カスパーゼ 11 が下流の細胞を活性化できることが研究でわかっています。 ギャップジャンクション チャネルタンパク質 パネキシン-1はK+流出を促進し、K+流出はNLRP3インフラマソームを活性化することができ、これによりカスパーゼ-1が活性化され、IL-1βおよびIL-18の成熟が促進されます。 [66].
2.2.3. GSDMDとパイロトーシスの関係
GSDMD (Gsdermin D) は、484 個のアミノ酸残基フラグメントから構成されるタンパク質であり、Gasdermin タンパク質ファミリーのメンバーです。このファミリーには主に 5 つのメンバーがあり、DFNA5、DFNB59、GSDMA、GSDMB、GSDMCD です。これら 5 つのタンパク質のアミノ酸構造は非常に類似しており、これらのタンパク質には N 末端と C 末端の 2 つの空間ドメインが含まれています。 [67]。 GSDMD タンパク質は長鎖アミノです 酸性タンパク質、N 末端ドメインと C 末端ドメインの間の構造は比較的緩やかです。 GSDMD は通常、次の位置で切断されます。 アミノ酸配列 275、N 末端ドメインと C 末端ドメインを露出します。で 哺乳類細胞GSDMD の単純な N 末端構造は、細胞のピロトーシスを制御できます。したがって、GSDMD は多くの場合 N 末端ドメインを発現しますが、すべての GSDMD タンパク質を発現できるわけではありません。 [68], [69]。しかし、研究により、GSDMDのN末端ドメインが培養のために大腸菌に導入されると、強い毒性効果が生じることが示されています。対照的に、無傷の GSDMD および GSDMD の C 末端ドメインは比較的低い毒性を示しました。 [70]。細胞内 GSDMD の N 末端構造は、損傷因子の侵入に伴って徐々に細胞膜に移行し、細胞膜上でホスホサルコシナーゼと特異的に結合し、 ホスファチジルセリン 細胞膜上で生物学的効果を生み出す [71]。原子力などの装置を使用する 電子顕微鏡法 L. Sborgi は、低温電子顕微鏡法により、GSMDD の N 末端構造がバイオフィルムと結合して、バイオフィルム上に中空の環状ポリマーを形成できることを観察しました。このポリマーはバイオフィルム細孔の形成を誘導し、これにより、内因性損傷が発生したときに GSDMD が細胞のピロトーシスを誘導します。外部の場合 病原体 侵入すると、病原体の細胞表面に吸着し、細胞を溶解することで外部の有害物質から体を守ることができます。 [72]。 GSDMD は、カスパーゼ-1/4/5/11 タンパク質の下流に特異的に結合する物質です。 シグナル伝達 [73]。カスパーゼ-1 は、さまざまなタンパク質との複合体によって生成されるインフラマソーム応答を通じて、下流の IL-1β および IL-18 の細胞膜からの放出を促進します。活性化されたカスパーゼ-1はGSDMDタンパク質を切断し、GSDMDを活性ペプチドに変換します。 GSDMDの分解生成物は細胞膜に特異的に結合し、細胞外水分子などの成分が細孔から細胞膜内に侵入し、細胞を膨張させ、最終的には破裂させます。 [74].
2.3.非コードRNAはISにおけるパイロトーシス遺伝子の発現に関与する
近年では、パイロトーシスの発生に関わる遺伝子レベルでの制御機構も完全に解明されてきました。現代の分子生物学の研究では、ゲノムの 98% がタンパク質のコード化に関与していないことが示されています。長鎖コーディング RNA (lncRNA) は、タンパク質をコードする機能を持たない RNA であり、長さが 200 ヌクレオチドを超えることからそのように名付けられました。 [75], [76]。ただし、 lncRNA タンパク質をコードする機能はありませんが、体内で最も発現され、転写において最も保存された遺伝子です。体内の重要な病理学的および生理学的プロセス、特に再灌流傷害関連疾患に関与し、重要な役割を果たしています。 [77]。研究により、ミクログリアでは、lncRNA-H19 の発現レベルが再灌流の期間と正の相関があることが示されています。 [78]。 LncRNA-H19 の過剰発現は、下流シグナル伝達分子 NLRP3/6 の発現を促進し、GSMDD を開始するときに炎症の役割を果たします。 [79], [80], [81]。 lncRNA-H19 はカスパーゼタンパク質ファミリーのメンバーを活性化することができます [82]ミトコンドリア機能不全を引き起こし、インフラマソームの活性化に関与し、神経損傷を引き起こします。 [83]。分子構造の制御に加えて、lncRNA-H19 はさらに多くの転写因子を動員して mRNA 転写プロセスを制御することもできます。 [84]。さらに、lncRNA-H19 は、 核輸送 転写因子のプロセスを制御し、それによってより多くの標的遺伝子の特異的発現を増加させ、炎症カスケードネットワークを生成します。 虚血 発生する [84]。したがって、lncRNA-H19 は、次のような場合に強い危険信号となります。 シリ H19 の阻害が起こる可能性があります。 処理 虚血再灌流傷害用 [84]。虚血再灌流傷害を引き起こす可能性のある AIM2 インフラマソーム媒介ピロトーシスでは、lincRNA MEG3/miR-485/AIM2 軸が CIRI 中にカスパーゼ 1 シグナル伝達を活性化することでパイロトーシスを促進するため、この軸は虚血再灌流障害の効果的な治療標的となる可能性があります。 は [84]。のために 免疫調節 ミクログリアの研究、Wang et al. LncRNA-Fendrr が ユビキチン化 HERC2 を介した NLRC4 タンパク質の分解とミクログリアのパイロトーシスの制御 [85]。張ら。 lncRNA NEAT1/miR-22–3p 軸がピロトーシスを抑制し、CIRI 損傷を軽減することを発見 [86]。 IS後のin vitro酸素グルコース欠乏(OGD)損傷実験では、OGDがNOD様受容体タンパク質3(NLRP3)の発現を増加させ、NSCのパイロトーシス死を誘導し、NSCは回復したことが示された。 高圧酸素療法。上方制御された lncRNA-H19 は miR-423-5p の分子スポンジとして機能し、OGD 後に NLRP3 を標的にして神経幹細胞 (NSC) ピロトーシスを誘導します。したがって、高圧酸素療法は、lncRNA-H19/miR-423–5p/NLRP3 軸を阻害することにより、NSC をピロトーシスから保護することが確認されました。 [87]。 miR-21は、 マイクロRNA (miRNA) パイロトーシスの発生と密接に関係しています。研究により、miR-21 は、炎症性フィードバック経路を介した NF-κB シグナル伝達を介して、NLRP3 タンパク質を特異的に制御し、NLRP3 タンパク質を活性化し、NLRP3 分泌を刺激し、マクロファージ内で発現できることが示されています。 [88]。 NLRP3 の活性化に加えて、miR-21 は下流のカスパーゼ-1 の活性化を調節し、IL-1β の分泌を促進し、パイロトーシスの発生を促進することもできます。 miR-21 が欠損すると、NLRP3 によって調節されるカスパーゼ 1 経路の発現が大幅に減少します。したがって、miR-21 の発現レベルもパイロトーシスの発生を制御する重要な要素となります。 [89], [90]。最新の分子生物学の研究では、miR-214–3p にはカスパーゼ 1 の結合部位があり、カスパーゼ 1 の発生に特異的に結合することで炎症反応を制御し、ピロトーシスの発生率を増加させることが示されています。 [91].
パイロトーシスの分子機構は次のように要約されています。 図1.

図1。ピロトーシスの分子機構 (ピロトーシスの機構には、標準的なインフラマソーム経路と非標準的なインフラマソーム経路が含まれます。標準的なインフラマソーム経路は、DAMP または PAMP によって引き起こされます。非標準的なインフラマソーム経路は、細胞外グラム陰性菌の LPS によって引き起こされます)トリガー後、一連の分子間相互作用を通じてパイロトーシスがトリガーされます。 TLR:トール様受容体。 GSDM: グスデルミン。 NLRP3: ヌクレオチド結合オリゴマー化ドメイン様受容体 pyrin ドメイン 3個を含む。 DAMP: 損傷に関連する分子パターン。 PAMP: 病原体関連分子パターン)。
2.4.パイロトーシスと他の細胞損傷との関係
脳虚血再灌流障害の発生には多くのメカニズムが関与しています。既知のメカニズムは、炎症反応、カルシウムチャネル障害、ミトコンドリア機能障害、オートファジーなどのさまざまなメカニズムに関連しています。パイロトーシスは脳虚血再灌流損傷のあらゆる側面に関与しており、他の損傷と密接に関連しています。
パイロトーシスとアポトーシスの発生はどちらも細胞死に属しますが、根本的に異なります。アポトーシスは細胞死の生理学的方法ですが、パイロトーシスは身体損傷後の病理学的細胞死に関連することがよくあります。 [92]。オートファジーとは、世界に広く存在するオートファジー現象の一種です。 真核細胞。オートファジーは一般に、シャペロン媒介オートファジー、マイクロオートファジー、マクロオートファジーの3種類に分類されます。現在、マクロオートファジーについては多くの研究が行われています。 [93]。オートファジーは、細胞の正常な代謝プロセスに不可欠な部分です。オートファジーが阻害されると、組織、器官、または細胞に有害な物質が蓄積する可能性があります。オートファジーが過剰に活性化すると、重要な細胞内小器官や必須タンパク質が破壊され、オートファジーアポトーシスが引き起こされます。 [94].
パイロトーシスとオートファジーは密接に関連しています。通常の状況下では、生体内でのオートファジーとパイロトーシスの発生は動的平衡状態にあります。体が外部刺激に対して炎症反応を起こすと、オートファジーとパイロトーシスのバランスが崩れます。 [95]。たとえば、NF-κB は シグナル伝達経路 多くのシグナル伝達に関与します 信号経路 シグナル伝達を達成します。この研究では、NF-κB遺伝子をノックアウトしたISマウスでは、 mTOR 活性が阻害され、オートファジーレベルが強化されました。 NF-κB のシグナル伝達活性化経路は、NF-κB 誘導キナーゼを活性化する多数の刺激であり、IκKα、IκKB、および IκKγ 三量体を活性化し、その結果 IκB のリン酸化と分解が起こり、最終的に NF-κB シグナル伝達経路が活性化されます。 。活性化後、NF-κB は細胞に入り、対応する DNA 受容体に特異的に結合し、それによって炎症因子の転写を刺激し、炎症因子 TNF-α、IL-1β、IL-6 の大量発現を促進します。研究により、NF-κB シグナル伝達経路が、パイロトーシスを誘導するための重要なパイロトーシスタンパク質 NLRP3 の転写前および転写後レベルの活性化の調節に関与していることが示されています。豊富に発現する炎症因子は、体内で炎症カスケードを誘発し、ピロトーシスを誘発し、さらに炎症反応を拡大させ、それによって脳損傷を悪化させる可能性があります。 [96], [97], [98].
2.5. ISに関与するパイロトーシスと炎症のメカニズム
炎症カスケードはISのさまざまな段階で存在します [99]。 ISの初期段階では、脳内の血流が遅くなり、 好中球 に従う 内皮細胞 虚血血管の損傷が起こり、初期の急性炎症反応が始まります。 [100]。脳虚血後に再潅流が起こると、外因性炎症因子、炎症細胞、炎症因子が血液脳関門を通過し、再潅流後の虚血による副次的な影響が生じ、多数の酸化因子や炎症因子が放出されます。 フリーラジカル。このとき、ミクログリアは 脳組織 徐々に大量に活性化され、より多くの量が生成されます。 炎症メディエーター。このタイプの炎症性メディエーターは、脳組織の内皮細胞を過剰に活性化し、大量の炎症性メディエーターを生成する可能性があります。 組織因子 アミノ酸の毒性が蓄積されます。これにより、酸化因子、フリーラジカル、および 一酸化炭素、NF-κB、Toll 様受容体、Nod 様受容体 (NLR) などの炎症反応関連シグナル伝達経路を活性化します。 [101], [102]。たとえば、NLR に関しては、研究により、NLR には 23 のファミリー メンバーが存在し、主に細胞質で発現され、体の自然免疫応答において重要な役割を果たしていることが示されています。 NLR ファミリーのメンバーである Nalp1、Nalp3、Nalp5、および Ipaf は、 アダプタータンパク質 ASC、炎症反応のカスケードを開始し、ピロトーシスを媒介します [103], [104]。 NLRP3 は炎症因子の重要な構成要素です。 NLRP3 には、PYD、NACHT、および LRR の 3 つのドメインが含まれており、LRR-NACHT-PYD: PYD-CARD: CARD-CARDC で表すことができます。スペース ドメインです。内因性損傷のさまざまなシグナルに反応するため、NLRP3 インフラマソームの活性化がピロトーシスの主なタイプであると考えられています。 [105], [106], [107].
インフラマソームは、体が感染または細胞損傷のシグナルを受け取った後に組み立てられるタンパク質複合体であり、プロカスパーゼ-1 の補充と活性化のプラットフォームとして機能します。プロモータータンパク質 NLRP3 は、インフラマソームアダプター分子 ASC を使用したオリゴマー化を通じて、大量のプロカスパーゼ 1 を動員できます。 [108]。プロカスパーゼ-1 はカスパーゼ-1 の前駆体であり、活性型カスパーゼ-1 は細胞を急速に溶解して細胞外 IL-1β および IL-18 を活性化して放出する役割を担っており、これにより炎症反応がさらに悪化します。したがって、パイロトーシスの発生は、炎症反応の出現およびインフラマソームの形成と密接に関連しています。脳虚血再灌流損傷の発生は、パイロトーシスと炎症反応の間の相互作用とより密接に関連しています [109], [110]。特に中枢神経系では、アストロサイトはミクログリアの活性化と増殖を誘導し、多数の炎症性メディエーターを生成します。これらの炎症性メディエーターは、内皮細胞を活性化してさまざまな組織因子を生成し、毒性を高める可能性があります。 興奮性アミノ酸のリリースを促進します。 一酸化窒素 そして酸素フリーラジカル。上記の物質はさらに、NF-κB や JNK2/STAT3 などの複数の炎症性シグナル伝達経路の活性化をもたらし、NLRP1 や NLRP3 などのインフラマソームの構築を促進します。活性化されたカスパーゼ-1は細胞のピロトーシスを誘発し、炎症反応を拡大し、脳虚血再灌流傷害を悪化させます [111], [112]。いくつかの研究では、野生型マウスと比較して、AIM2の脳内のAIM2およびIL-1βのレベルが低いことが判明しました。 ノックアウトマウス 脳虚血再灌流が大幅に減少した後、梗塞体積は大幅に減少し、 神経機能 スコアも大幅に向上しました。 AIM2 インフラマソームの活性化を阻害すると、ピロトーシスの発生をある程度抑制できるため、虚血性脳損傷が軽減されます。 [113]。したがって、パイロトーシスは脳虚血再灌流障害の過程で重要な役割を果たします。 NLRP3 インフラマソームはパイロトーシス経路の重要なタンパク質であり、その発現を阻害すると、下流のパイロトーシス経路関連タンパク質の発現も同時に阻害され、炎症反応が制限され、脳虚血再灌流傷害が緩和されます。
2.6.パイロトーシスとミトコンドリア損傷はIS後の酸化ストレス損傷を媒介する
ミトコンドリアは主に細胞の好気呼吸に関与しており、好気呼吸が起こる場所です。ミトコンドリアは、細胞間の正常な信号伝達と神経細胞の信号伝達のためのイオン化された基盤を提供します。 [114]。ミトコンドリアの損傷は主に異常によって引き起こされます ミトコンドリア代謝 そして酸化ダメージ [115]。研究により、脳虚血再灌流損傷はミトコンドリア機能不全と密接に関連していることが示されています。 [116]。酸化ストレスは酸化損傷発生の鍵であり、パイロトーシスと密接に関係しています [117], [118]。酸化ストレスは、脳組織損傷、特に脳虚血や脳虚血で発現する可能性が高くなります。 再灌流傷害 脳虚血後 [119]。体が外部からの有害物質の攻撃を受けると、細胞は大量の有害物質を分泌します。 ROS そして 活性炭酸化と抗酸化のバランスを崩し、一連の酸化損傷反応を引き起こします。 [120]。 ROS のほとんどはミトコンドリアによって分泌されます [121]。外部刺激または内因性損傷によって刺激されると、体は高レベルの ROS を誘導し、インフラマソームタンパク質複合体を活性化することにより、NLRP3 へのヌクレオチドの結合を刺激します。 ROS が過剰に活性化すると、体内のシグナル伝達経路の正常な順序が破壊され、その結果、細胞内の遺伝物質、タンパク質、およびさまざまな細胞小器官 (ミトコンドリアを含む) が破壊されます。細胞内のミトコンドリアなどのさまざまな細胞小器官が損傷すると、細胞の栄養や代謝に異常が生じ、活性酸素が細胞の損傷をさらに悪化させる可能性があります。 血管内皮 の撹乱を誘発します。 微小循環 脳の中で。脳の血液脳関門の透過性障害は、 細胞接着分子、それによって脳損傷を悪化させる [122]。その間、 酸化ストレス 細胞が徐々に増加することを特徴とする細胞死モードを示すようになります。 細胞膜透過性 そして細胞内容物の放出 – パイロトーシス [120]。したがって、ピロトーシスとミトコンドリア損傷の両方が脳虚血再灌流損傷の病理学的過程に関与しています。ミトコンドリア運動関連タンパク質Drp1は、ミトコンドリアが正常な生理機能を維持するために必要な重要なタンパク質であることが報告されています。 [123]。 Drp1 の発現が増加すると、 ミトコンドリア分裂 パイロトーシスのレベルを処理して低下させ、それによって損傷の発生を遅らせます。 [124]。身体がさまざまな内因性および外因性の損傷刺激を受けると、体内のシグナル伝達経路の制御プロセスが変化します。 Drp1の分泌低下、ミトコンドリアの損傷、ミトコンドリアの機能不全、アポトーシスやパイロトーシスの増加として現れ、体内のさまざまな疾患を引き起こします。 [125]。最近の研究では、ミトコンドリア機能不全が脳虚血/再灌流損傷中に NLRP3 インフラマソーム活性化を誘導することが示されています。 [126]。ミトコンドリア 脱共役タンパク質 2 (UCP2) 欠損は CIRI 後の脳損傷を悪化させ、新しい研究では、UCP2 欠損が高血糖誘発性 CIRI 増悪後の in vitro および in vivo での NLRP3 インフラマソーム活性化を促進することが示されています。 UCP2 は、高血糖によって誘発される CIRI の悪化に対する潜在的な治療標的となる可能性があります。 UCP2欠損はまた、in vitroおよびin vivoでニューロンにおけるNLRP3インフラマソーム活性化とROS産生を増強した [127]. アディポネクチン は、広範な抗酸化作用と抗炎症作用を持つ脂肪由来のホルモンです。アディポネクチンペプチドはAMPK/GSK-3βを調節することにより脳虚血再灌流障害後の酸化ストレスとNLRP3インフラマソーム活性化を軽減する [128]。したがって、ISでは、パイロトーシスの発生とミトコンドリア損傷の間に密接な関係があり、両者の間には密接な相互作用があります。
2.7.酸化ストレス/ニトロソ化ストレスはIS後のパイロトーシスを媒介する
酸化/ニトロソ化ストレスと 神経炎症 鍵です 病理学的プロセス 虚血性脳卒中における脳虚血再灌流損傷、媒介ニューロン損傷、血液脳関門損傷および出血性変化の研究 [129], [130]。一酸化窒素(NO)と ペルオキシ亜硝酸塩 (ONOO-) は CIRI の典型的な RNS です [130], [131]。の異性体は3つあります 一酸化窒素合成酵素: 内皮一酸化窒素合成酵素 (eNOS) 低濃度の NO を生成し、生理学的機能を持ちます。その間 神経の一酸化窒素合成酵素 (nNOS) および 誘導性一酸化窒素合成酵素 (iNOS) 高濃度の NO を生成し、炎症を誘発し、血液脳関門の透過性を高めます。 [132]。 ROS/RNS の過剰生産はストレスの多い微環境を作り出し、一連の細胞シグナル伝達カスケードを引き起こし、炎症、血液脳関門の透過性亢進、 脳浮腫 そして神経細胞死 [133], [134]。ペルオキシナイトライト媒介 DNA鎖 を破壊してアクティブ化します リボザイム ポリ(ADP-リボース)シンターゼ(PARS)、ポリ(ADP-リボース)ポリメラーゼ(PARP)またはポリ(ADPリボース)としても知られています。 転移酵素 (pADPRT)。 ONOO - PARP を直接アクティブ化できます [135]、そしてインビボおよびインビトロ実験は、虚血性脳損傷におけるONOO-/PARPシグナル伝達経路の役割を実証した。インビボ実験では、虚血性脳卒中の動物モデルにおいて、NOS欠損マウスではPARP活性化が少ないことが示された。インビトロ実験では、ONOOドナーは培養C6においてPARP活性化を有意に誘導するが、NOドナーは誘導しないことが示された 神経膠腫細胞. 遺伝子破壊 またはPARPのサイレンシングにより大幅に減少します 脳梗塞の大きさ、軽減します 神経毒性、神経血管ユニットを保護し、神経学的転帰を改善します。 [136], [137], [138]。研究により、ROS/RNS が CIRI 中のインフラマソームの活性化因子である可能性があることが判明しました。 [139]。インフラマソームはミクログリアの細胞質にある複数のタンパク質複合体であり、その中で NOD 様受容体 3 (NLRP3) が最も広く研究されています。 NLRP3 はカスパーゼ-1 を誘発して活性化し、IL-1β と IL-18 を活性化して体内に放出します。 細胞外空間、炎症の進行を促進する [140]。 in vivo実験では、NLPR3ノックアウトマウスの脳梗塞サイズとBBB損傷が野生型マウスよりも低いことが示された。さらなる実験により、NLRP3がIL-1βの放出を媒介し、脳微小血管内皮細胞の透過性を高めることができることが示されました。 [141]。核因子E2関連 係数2 (Nrf2) は細胞の抗酸化反応を調節し、Nrf2 は酸素およびグルコース欠乏条件下で BV2 ミクログリアにおける ROS 媒介 NLRP3 産生を阻害します。 [142]。酸化/ニトロソ化ストレスがペルオキシ亜硝酸塩の形成を誘導し、これがカスパーゼ1/インフラマソーム活性化の重要なトリガーである可能性があることが判明しました。 [143]。したがって、ROS/RNS 媒介インフラマソームは、虚血性脳損傷の潜在的な治療標的となる可能性があります。
さらに、ROS/RNS は、Toll 様受容体の活性化を仲介します。現在の研究では、Toll 様受容体 (TLR) が先天性受容体として広く研究されていることが示されています。 免疫受容体、TLR4/2 は大脳分野でさらに研究されています。 虚血性損傷. TLR4 IL-1β、MMP-9、iNOS、COX-2の発現を媒介し、酸化ストレス誘発性の脳損傷を悪化させる可能性がある [144]。 TLR4阻害剤の適用 E5564 を再生できることを示しました。 神経保護的な ミクログリアの活性化とROSの産生を阻害することによる役割 [145]。 IS後、TLR4欠損マウスの梗塞サイズは野生型マウスの梗塞サイズよりも有意に小さかった [146]。 TLR4はポストISにも関与 神経新生. 陽電子放出断層撮影 研究では、TLR4欠損マウスがISにおける神経新生を増強し、炎症反応を抑制することが示されています。 [147]。さらに、in vivo 実験研究では、TLR2/4/NF-κB シグナル伝達経路の阻害が、酸化ストレス、炎症反応の調節、虚血性脳組織の保護に一定の効果があることを示しています。 [148]。したがって、TLR4/2 は虚血性脳損傷の治療標的となる可能性があります。
3. パイロトーシスとISの関係
大脳の病理学的メカニズム 虚血 虚血再灌流後の傷害は複雑です。このプロセスに関与するメカニズムには、炎症、酸化ストレス、オートファジー、ミトコンドリア機能不全、カルシウム過剰、プログラム細胞死などが含まれます。 [149], [150], [151]。プログラムされた細胞死が脳虚血および虚血再灌流障害の病理学的過程において重要な役割を果たしており、上記の機構、特にパイロトーシスと絡み合って密接に関連していることが、多くの研究で示されている。 [152], [153]。プログラムされた細胞死は、ニューロンが関与する虚血性脳卒中の主要なイベントです。 ミクログリア、アストロサイト、 血管内皮細胞、アポトーシスなど [154]、オートファジー [155]、プログラムされた壊死または壊死症 [156], フェロトーシス [157]、およびパイロトーシス [158]。特にパイロトーシスでは、ラット脳虚血モデルにおいて NLRP1、ASC、カスパーゼ-1、GSDMD などのパイロトーシス関連タンパク質の発現が増加しており、NLRP1 を阻害すると炎症や脳損傷を軽減することができます。 虚血性損傷。さらに、慢性脳虚血の場合、細胞は DAMP や PAMP、そして一連の複雑な分子反応を引き起こします。このとき、ミクログリアやアストロサイトなどの頭蓋内神経免疫炎症細胞が増殖して活性化し、神経血管単位にさらに損傷を与えるようです。 [159], [160], [161].
3.1.パイロトーシスとニューロン
IS後は短時間で虚血中心部に壊死が起こり、死んだ細胞が次のような危険信号を発します。 HMGB1タンパク質, ヒートショックプロテイン, ペルオキシダーゼ これらの危険信号分子はファミリータンパク質などに結合します。 パターン認識受容体、 形状 インフラマソーム、自然免疫反応を開始し、神経細胞死を引き起こす [162], [163]。研究により、脳虚血が NLRP1 と NLRP1 の高発現を引き起こす可能性があることが確認されています。 NLRP3 虚血性脳組織およびニューロンでは、NLRP1 の活性化は主にニューロンに存在します。 [164]。カスパーゼ-1 阻害剤または免疫グロブリン製剤を使用すると、一次皮質ニューロンにおける NLRP1 および NLRP3 の発現が減弱し、神経細胞のサイズが縮小する可能性があります。 脳梗塞、そしてそのメカニズムはNF-κBとMAPK経路の活性化の阻害に関連している可能性があります。 [164]。 IS マウスでは、Li et al. [165] 脳虚血後3日目に、神経細胞質、核、および神経細胞の超微細構造損傷が発見された。 ミトコンドリア膜 が起こり、Caspase-1、GSDMD、IL-1βの発現が有意に増加しました。カスパーゼ-1阻害剤 Vx765 ピロトーシスを抑制し、虚血領域のニューロンの生存を促進し、症状を改善することができます。 脳機能障害 マウスで。リャンら。 [166] は、長い非コーディング RNA 母性発現遺伝子 3 (MEG3) が、AIM2/カスパーゼ-1 経路を活性化することによってピロトーシスと炎症反応を促進し、脳虚血再灌流傷害を引き起こすことを発見しました。 MEG3 遺伝子をノックアウトすると、次のような発現が阻害されます。 AIM2、カスパーゼ-1、GSMDD およびその他のタンパク質が含まれており、虚血性脳損傷を軽減します。これは、MEG3 が虚血性脳卒中に対する効果的な治療標的である可能性があることを示唆しています。
3.2.パイロトーシスとアストロサイト
アストロサイトが最も豊富です グリア細胞 の中に 中枢神経系 血液脳関門の形成、神経代謝の調節、細胞間コミュニケーションの安定化に関与しています。 [167]。 IS後の顕著な病理学的変化は反応性である アストログリア症 虚血性損傷後の早期に神経の可塑性を促進するグリア瘢痕化 [168]。一般に信じられているのは、 海馬 虚血であり、 低酸素症、 抗酸化力 海馬の機能が低下し、多くの炎症因子が放出され、海馬へのダメージが悪化する可能性があります。 [169]。局所脳虚血から 3 時間後 SDラット、IL-1β様免疫反応性陽性細胞が虚血領域に出現し、主に不活化星状細胞が出現した。虚血後 2 か月まで、陽性細胞は虚血半球、特に壊死巣の周囲で検出され、活性化の程度が高くなります。 [170]。活性化されたアストロサイトは、 腫瘍壊死因子、インターロイキン、成長因子、その他の炎症因子または神経毒性メディエーターにより、神経損傷が引き起こされます。 [171]。 NLRP2 は主にアストロ サイトで発現されますが、ニューロンやミクログリアではほとんど発現されません。さらに、NLRP2の発現は、マウス脳虚血モデルおよび星状細胞の酸素およびグルコース欠乏後に有意に上方制御され、NLRP2をサイレンシングすることで、酸素およびグルコース欠乏によって誘発されるピロトーシスを軽減できる可能性がある。 [172]。別の研究では、脳虚血の in vitro モデルにおいて、酸素グルコース欠乏により NLRP3、ASC、カスパーゼ-1、IL-1β、および IL-18 が増加し、星状細胞の生存が減少することがわかりました。 ヒスピドゥリン から単離されたケトン化合物です。 漢方薬。 in vitro および in vivo 実験では、ヒスピジュリンが NLRP3 媒介ピロトーシスを阻害し、脳に保護効果を及ぼすことができ、そのメカニズムは AMPK/GSK-3β の活性化に関連していることが示されています。 シグナル伝達経路 [173]。孟ら。 [174] NLRP6 の発現は、ラットの脳虚血再灌流後 48 時間でピークに達することを発見しました。アストロサイトで酸素とグルコースが欠乏すると、NLRP6 とその活性化産物の産生が増加し、NLRP6 をサイレンシングすると ASC とカスパーゼ 1 が減少し、炎症因子の放出が減少し、ニューロンの活動が増加します。 [175]。したがって、星状細胞におけるインフラマソーム活性化を標的とした介入は、IS の治療に新しいアイデアを提供する可能性があります。
3.3.パイロトーシスとミクログリア
ミクログリアは先天的に存在する 免疫細胞 中枢神経系と虚血性脳卒中後の最も初期に活性化された細胞の [176]。脳虚血損傷の急性期では、ミクログリアが急速に病変部位に移動し、炎症因子や細胞傷害性物質を分泌し、組織損傷を悪化させます。 [177]。慢性期では、ミクログリアは抗炎症性サイトカインと成長因子を生成して炎症を促進します。 組織修復 そして改造 [178]。研究により、ミクログリアのピロトーシスが虚血性脳損傷において重要な役割を果たしていることがわかっています。 NLRC4は、ミクログリアが酸素とグルコースを3時間欠乏させたときに有意に増加した最初のインフラマソームであったが、NLRP1、NLRP3、およびAIM2は、酸素グルコース欠乏の6時間後まで有意に増加しなかった。 NLRC4 をサイレンシングすると、GSDMD、IL-1β、IL-18 の産生が減少し、ミクログリアのピロトーシスが抑制される可能性があります。 [179]。徐ら。 [180] は、ドライバー受容体 1 (TREM-1) が次のように発現していることを報告しました。 骨髄細胞 脳虚血再灌流後のミクログリアでは、NLRP3/カスパーゼ-1媒介ピロトーシス経路が活性化され、神経炎症反応が誘導される可能性があります。 TREM-1 を阻害すると、ミクログリアのパイロトーシスと神経損傷が軽減されます。リーら。 [181] 環状グアノシン一リン酸アデノシン合成酵素 (cGAS) の発現が脳虚血後に上方制御され、AIM2 インフラマソームを活性化してミクログリアのピロトーシスを誘導することを確認しました。 cGAS アンタゴニスト A151 は、AIM2 活性化とミクログリアのピロトーシスを阻害し、脳梗塞体積を大幅に減少させ、神経損傷を軽減します。上記の研究は、ミクログリアのパイロトーシスとその媒介性神経炎症反応が虚血性脳損傷を引き起こす重要なメカニズムである可能性があることを示しています。ミクログリアの神経毒性作用を抑制することは、虚血性脳卒中治療の新しい戦略となる可能性があります。
3.4.ピロトーシスと内皮細胞
内皮細胞は血液脳関門の最初の関門を構成し、血液脳関門の足場を提供します。 クローディン, 接着分子 そして 細胞外マトリックス [182]。脳虚血中、免疫炎症反応と酸化ストレスにより損傷が生じる可能性があります。 内皮細胞 血液脳関門の完全性を破壊し、血管原性浮腫、出血性変形、死亡率の増加を引き起こします。 [183], [184]。脳虚血後、NLRP3 はニューロン、ミクログリア、血管内皮細胞で上方制御され、NLRP3 遺伝子をサイレンシングすると、脳虚血マウスの脳梗塞の量を減らすことができます。 中大脳動脈 虚血と血液脳関門の透過性の低下 [185]。王ら。 [186] 脳虚血が微小血管内皮細胞のピロトーシスを誘発し、虚血再灌流傷害を悪化させる可能性があることを確認しました。彼らはまた、ペルオキシソーム増殖因子活性化受容体ガンマコアクチベーター 1α (PGC-1α) の活性化により、パイロトーシス関連タンパク質の発現が大幅に減少し、ZO-1 および ZO-1 の発現が増加することも発見しました。 オクルディン 血液脳関門の完全性を保護するために、脳微小血管内皮細胞内のタンパク質。さらに、脳微小血管内皮細胞のピロトーシスにより、IS後の出血の可能性が高まり、次のような重篤な合併症を引き起こす可能性があることが研究で判明しています。 脳出血 [187].
脳虚血とパイロトーシスの関連性は次のように要約されています。 図2.

図2。 IS後の血管神経細胞におけるピロトーシスのメカニズムのまとめ [血管神経細胞(ミクログリア、アストロサイト、ニューロン、神経細胞からなる)におけるピロトーシスの分子機構 血管内皮細胞、周皮細胞)IS後。 IRF: インターフェロン調節因子; IFN: インターフェロン; CASP: カスパーゼ。 GPX4: グルタチオンペルオキシダーゼ 4]。
4. IS治療におけるパイロトーシス阻害剤の可能性
近年、ISにおけるパイロトーシス阻害剤の研究が大きな注目を集めています。リャンら。 [188] カスパーゼ 1 阻害剤 VX-765 が、カスパーゼ 1、ASC、GSDMD、IL-1β のレベルをアップレギュレートしながら、カスパーゼ 1、ASC、GSDMD および IL-1β を減少させることができることを発見しました。 密着結合タンパク質 そして メタロプロテイナーゼの組織阻害剤、血液脳関門の完全性を保護します。さらに、VX-765 はミクログリアの M1 型から M2 型への変換を促進し、ミクログリアによる炎症反応を軽減し、神経保護効果を発揮します。 [189], [190]。 MCC950 は選択的 NLRP3 阻害剤です。研究では、MCC950 が虚血周縁部における NLRP3、カスパーゼ-1、および IL-1β の発現を減少させることができ、マウスの局所脳虚血に対して保護効果があることが判明しました。 [191]。研究によると、いくつかの マイクロRNA 脳虚血モデルにおけるピロトーシスを阻害し、神経保護の役割を果たす可能性がある [192]。 LP17は阻害する可能性があります 骨髄細胞 受容体-1をトリガーし、それによって酸化ストレスとパイロトーシスを抑制し、脳虚血誘発性の神経損傷を軽減します。 [192]. ヒスチジン アデニル酸活性化プロテインキナーゼ/グリコーゲンシンターゼキナーゼ-3βシグナル伝達経路を調節することでNLRP3媒介ピロトーシスを阻害し、それによって抗CIRI神経保護効果を発揮する可能性がある [193]。低密度リポタンパク質受容体は、CIRI後のNLRP3媒介神経ピロトーシスを阻害する可能性がある [194]。脂質アルコール媒介ペルオキシソーム増殖因子活性化受容体α-グルタミン酸オキサロ酢酸トランスアミナーゼ1軸は、IS後の内皮細胞のパイロトーシスを阻害し、虚血性脳損傷を改善する可能性がある [195]。 CHRFAM7Aの過剰発現は、NLRP3/カスパーゼ-1経路依存性のミクログリアのピロトーシスを阻害し、CIRIを軽減する可能性がある [196]。 TP53 誘導性の解糖およびアポトーシス制御因子は、脳虚血誘導性のミクログリアのピロトーシスおよび虚血性脳損傷を軽減する可能性がある [197]。加えて、 エクソソーム 低酸素骨髄由来 間葉系幹細胞 ミクログリアのM1/M2表現型を調節して、CIRI誘発性ニューロンパイロトーシスを緩和する可能性がある [198]。低酸素前処理済み 嗅粘膜 間葉系幹細胞は、低酸素誘導因子-1αを活性化することで脳虚血によって引き起こされるミクログリアのピロトーシスとアポトーシスを阻害し、それによってCIRIを低下させる可能性がある [199].
パイロトーシスの実行者である GSDMD は、IS 治療の理想的な分子標的です。しかし、GSDMD阻害剤に関する現在の研究はまだ初期段階にあり、虚血性脳卒中治療におけるGSDMD阻害剤の有効性を示す証拠は不足している。最近の研究では、一部の天然植物成分がパイロトーシス経路を調節することによって IS を治療できる可能性があることが判明しました。
5. 新しいパイロトーシス阻害剤としての天然物は、IS 治療の潜在的候補となる
天然物は、多成分、多標的、多チャネル、低毒性という利点により、脳梗塞/脳虚血再灌流の予防と治療に大きな利点があり、優れた応用の見通しを持っています。現在、脳梗塞/CIRIの予防と治療における天然物の研究は徐々に増加しており、さまざまな防御機構が関与しています。このセクションでは、安全で効果的で毒性が低い天然植物成分を迅速に探索するための理論的裏付けを提供するために、天然物、動物または細胞モデル、投与方法、投与量、治療時間、効果、および治療メカニズム(シグナル伝達経路)を要約しました。 、IS/CIRIの予防と治療のためにピロトーシスを制御します。
5.1.天然植物成分
天然植物成分の主な構造は次のとおりです。 図3.

図3。天然植物成分の主な構造。
5.1.1.ガストロジン (GAS)
GASはハーブ由来の多機能化合物であり、 ガストロディア・エラタ、さまざまな 薬理活性、抗酸化作用や抗炎症作用など。さらに、GAS は、カスパーゼ-3 切断、IL-18、IL-1β などのアポトーシス促進因子を調節することにより、脳虚血再灌流の症状を大幅に軽減することが示されています。いくつかの研究では、GAS が脳虚血損傷を改善し、脳虚血レベルを低下させることが示されています。 活性酸素種 そして 炎症性サイトカイン マウスで [200], [201], [202]。ルーら。 GASは、NLRP3/カスパーゼ-1軸を標的とすることにより、心臓微小血管虚血再灌流傷害に関連するピロトーシスを改善できることを発見した [203]。 GAS は、 神経機能 スコアを付けてサイズを縮小する 脳梗塞。一方、GAS は、NLRP3、炎症因子 (IL-1β、IL-18)、および切断型カスパーゼ 1 を下方制御することによりパイロトーシスを抑制しました。さらに、GAS は、lncRNA NEAT1/miR-22-3p 軸を調節することにより、虚血再灌流によって誘発される神経細胞の炎症を軽減しました。 GASはlncRNA NEAT1/miR-22-3p軸を調節することにより脳虚血再灌流障害を大幅に軽減した [204]。したがって、GAS はポテンシャルとして使用される可能性があります。 薬 CIRIの治療に。
5.1.2.タンシノーネIIA
タンシノーネIIA ~の主な有効成分です サルビア・ミルティオリザ。心血管疾患や心臓病の治療に広く使用されています。 脳血管疾患 抗酸化作用と血行改善作用があるためです。蔡ら。 [205] タンシノン IIA が NLRP3 炎症性シグナル伝達経路における IL-1β および IL-18 の発現を効果的に減少させ、その効果が用量依存性であることを発見しました。これは、タンシノン IIA が NLRP3 シグナル伝達経路の活性化を阻害し、それによって脳卒中後の BV-2 細胞に対する酸素-グルコース欠乏/再灌流の損傷を軽減することを示唆しています。
5.1.3.レオヌリン
レオヌリン で レオンリ ハーバ 慢性脳虚血ラットの学習能力と記憶能力を改善し、NLRP3の発現を減少させました。神経を保護するという目的は、抗炎症作用によって達成されるのではないかと推測されています。 NLRP3 の活性化により、カスパーゼ 1 前駆体のカスパーゼ 1 への変換が促進され、IL-1β および IL-18 の産生が促進され、それによって一連の炎症反応が引き起こされました。レオヌリンは脳組織の炎症反応を軽減し、細胞のピロトーシスを阻害することで神経学的欠損スコアを低下させ、良好な神経保護効果を有する可能性があります。 [206].
5.1.4.ペオニフロリン
ペオニフロリン ~の主成分に由来する シャクヤク radix redx。ペオニフロリンは、グルコシドの総グルコシドの主な活性成分です。 牡丹 (TGP)、優れた抗炎症作用および免疫調節作用もあります。 [207]。 TGPやペオニフロリンも、次のような様々な薬理活性を持っています。 鎮痙薬, 鎮痛剤、 そして 血管拡張 [208]。ズリフィヤ・エケムら。急性脳虚血再灌流後、脳内のミクログリアと神経ピロトーシスの活性化が明らかであり、NLRP3の活性化が増加し、カスパーゼ-1とIL-1βが上方制御されていることを発見しました。 [209]。ペオニフロリンは神経機能スコアを大幅に低下させ、大幅に低下しました 脳梗塞の大きさ、Iba1発現が大幅に下方制御され、ニューロンの状態が大幅に改善されました。さらに、NLRP3、カスパーゼ-1、およびIL-1βの発現は、ペオニフロリン介入後に有意に減少した。ペオニフロリンは、NLRP3 と Caspase-1 の活性化を阻害することにより、ミクログリアの活性化を低下させ、ニューロンのピロトーシスを阻害し、ACI を介した神経損傷を改善する可能性があることが示唆されています。 [209].
5.1.5.ブチルフタリド
L-ブチルフタリドは最初にクレソンの種子から単離され、後に人工的に合成できるようになりました。 ブチルフタリド ユニークな二重の役割を持ち、再構築するだけでなく、 微小循環 虚血再灌流を増加させ、それによって血管の完全性を保護します。 血管構造、血管の直径を回復し、虚血領域と周囲の血流数を増加させます。 微小血管だけでなく、ミトコンドリアを保護し、細胞死を減らすことで、ミトコンドリア構造の完全性を保護し、ミトコンドリア複合体酵素 IV の活性を改善し、ミトコンドリア ATP 酵素の活性を改善し、ミトコンドリア ATP 酵素の安定性を維持します。 ミトコンドリア膜 [210]。最新の研究では、ブチルフタリドが NLRP3 インフラマソームシグナル伝達経路を介して用量依存的に CIRI ラットのピロトーシスに影響を与える可能性があることが示されています。
5.1.6.ブレビスカピン
ブレビスカピン から抽出されます エリゲロン・ブレビスカプス (ヴァント。)手。 -マズ。ブレビスカピンは以下の混合物です。 スクテラリン スクテラリンを主成分とし、脳血管を拡張し、脳血管抵抗を軽減し、脳血流を増加させ、微小循環を改善し、抵抗力を改善する機能を有するスクテラリンを少量含む。 血小板凝集。最新の研究では、ブレビスカピンがNLRP3炎症細胞の活性化を有意に阻害したことが示されています。 海馬 CCI ラットのカスパーゼ 1、IL-6、IL-1β タンパク質の発現を下方制御し、カスパーゼ 3 タンパク質の活性化を阻害し、 ニューロンのアポトーシス。これは、ブレビスカピンが CCI ラットの認知機能を大幅に改善し、虚血ニューロンの病理学的損傷を軽減する可能性があり、そのメカニズムが脳組織における NLRP3 インフラマソーム活性化とパイロトーシス経路の阻害に関連している可能性があることを示唆しています。 [211].
5.1.7.レスベラトロール
レスベラトロール 非フラボノイドです ポリフェノール化合物。それは最初に根から分離されました。 ベラトラム グランディフロラム、レスベラトロールは700以上の植物で発見されています。研究では、レスベラトロールが脳梗塞における酸化ストレスの調節、炎症の抑制、脳の状態の改善に重要な役割を果たすことが示されています。 神経保護。レスベラトロールは、主にミクログリア NLRP3 インフラマソーム、カスパーゼ 1 および ZO-1 の制御を通じて、虚血再灌流脳組織のパイロトーシスを制御します。 [212].
5.1.8.サルビアノール酸 (SAFI)
最近の研究では、SAFI が神経学的欠損スコアを大幅に増加させ、梗塞体積を減少させ、組織学的損傷を軽減することが示されています。 大脳皮質 MCAO/R モデルでは神経細胞のアポトーシスが起こり、OGD モデルでは神経細胞の生存率が増加し、神経細胞のアポトーシスが減少します。 SAFIはまた、ミクログリアの分極パターンをM1様表現型からM2様表現型に改造し、NLRP3インフラマソームの活性化およびNLRP3インフラマソーム/発熱関連タンパク質の発現をin vitroおよびin vivoで阻害した。これは、SAFIがニューロンのアポトーシスを減少させ、ミクログリアの表現型をM1からM2に移行させ、ミクログリアのNLRP3インフラマソーム/パイロトーシス軸を阻害することによって神経保護効果を発揮する可能性があることを示唆しています。 [213].
5.1.9.ヒスピドゥリン
ヒスピドゥリン です フラボノイド さまざまな薬理学的特性を持つ化合物であり、多くの薬の主要な有効成分の 1 つです。 漢方薬。ヒスピドリンには、抗酸化物質、 抗真菌剤, 抗悪性腫瘍、抗骨粗鬆症、抗炎症、抗変異原性の特性。最新の研究では、ヒスピドゥリンが症状を改善することが示されました。 神経症状 IRI後のラットでは、梗塞サイズが減少し、 脳浮腫。機構的には、ヒスピドリンは、AMPK/GSK3β シグナル伝達経路を調節することで NLRP3 媒介ピロトーシスを阻害することにより、生体内および生体外で神経保護効果を発揮します。 [214].
5.1.10.アストラガロシド IV
黄耆基数 欠乏を補い、脾臓と胃を強化し、血液循環を促進し、血液循環を促進し、次の症状を治療します。 気の欠乏 そして血液欠乏症 [215]. アストラガロシド IV の主要な有効成分の 1 つです。 黄耆基数 そしてその モノマー の成分 レンゲ サポニン。レンゲの品質を検査するための基準としてよく使用されます。 [216]。アストラガロシド IV にはさまざまな種類があります。 生物活動、研究範囲には脳、肝臓、心臓、肺、腎臓、胃、腸、血管などの複数の臓器や組織が含まれます。 [217], [218], [219] 抗炎症、抗ウイルス、抗アポトーシス、促進などの生物学的活性があります。 細胞増殖、規制する ウサギの病気、血糖値を調節し、老化を遅らせ、がんを抑制します。 [217], [218], [219]。私たちの以前の研究では、アストラガロシド IV が IS の治療に重要な役割を果たす潜在的な神経保護剤であることが特定されました。例えば、血液脳関門を保護し、エネルギー代謝を改善し、神経細胞のアポトーシスを阻害し、炎症反応や酸化ストレスを阻害し、神経保護効果を発揮して脳虚血を改善することができます。 [220], [221], [222]。パイロトーシスに関しては、Tang et al. NLRP3 インフラマソームはラットの脳虚血再灌流中に活性化され、NLRP3 インフラマソームを阻害するか、その下流のカスパーゼ 1 を阻害すると、ラットの脳虚血再灌流の損傷が軽減される可能性があることを発見しました。 [223]。アストラガロシド IV は、神経障害スコアを低下させ、脳梗塞の容積を減少させ、脳組織における NLRP3、カスパーゼ-1、プロ IL-1β、IL-1β、プロ IL-18、および IL-18 のタンパク質レベルを低下させることができます。 、リン酸化されたNF-κBタンパク質の発現を阻害します。これは、アストラガロシド IV には抗 CIRI 効果があり、そのメカニズムは NF-κB の阻害に関連している可能性があることを示唆しています。 タンパク質のリン酸化 NLRP3 インフラマソーム活性化の阻害 [223]。リーら。また、アストラガロシド IV が新生仔ラットの低酸素虚血誘発性脳損傷を軽減し、MMP-9 媒介 NLRP3/カスパーゼ-1 シグナル伝達経路 [224].
5.1.11.オタネニンジンサポニン (PNS)
人参根と根茎 乾燥した根です 根茎 の パナックス 能登銀泉(バーク) FH Chen、血液、心血管、神経、免疫系に薬理活性を有する [225], [226]。 PNS は主な有効成分であり、さまざまな単量体サポニンを含み、Xuesaitong 注射剤などのその製剤は心血管疾患や脳血管疾患の予防と治療に広く使用されています。 [227]。私たちの以前の研究と既存の研究は、PNSが虚血性脳損傷に対してさまざまな薬理学的効果を持っていることを示しています。 抗血栓薬、抗炎症、抗酸化、脳神経細胞のアポトーシスの阻害、血液脳関門の損傷の改善 [228], [229], [230], [231]。これは、PNS が複数の標的を通じて脳虚血において保護的な役割を果たしている可能性があることを示唆しています。最近の研究では、PNS が NLRP3 インフラマソームの活性化を阻害し、選択的に促進することができることが判明しました。 マイトファジー ラットの脳虚血再灌流中。マイトファジーを阻害すると、NLRP3 インフラマソームに対する PNS の阻害効果が逆転する可能性があり、マイトファジーが脳虚血再灌流における NLRP3 インフラマソーム活性化に対する PNS の阻害効果を媒介することが示されました。さらに、ラットの脳虚血再灌流中にミトコンドリア内のPINK1およびParkinタンパク質のレベルが増加し、PNSは脳組織のミトコンドリア内のPINK1およびParkinタンパク質のレベルをさらに増加させる可能性がある。これは、PNS が PINK1/Parkin 経路を介して脳虚血再灌流におけるマイトファジーを促進する可能性があることを示唆しています。 [232]。要約すると、PNS は PINK1/Parkin 経路を通じてマイトファジーを選択的に促進し、脳虚血再灌流における NLRP3 インフラマソームの活性化を阻害し、脳虚血再灌流傷害を軽減します。
5.2.天然植物エキス
5.2.1.タオホンシウ煎じ薬 (THSWD)
THSWD は、血液循環を促進し、血液の循環を促進し、 瘀血、清朝の呉謙の「医学の金鏡」によって作成された、血液の停滞を除去し、新しい血液を再生します。現在の薬理学的研究により、THSWD の主な有効成分には次のものがあることが確認されています。 フェルラ酸, ベニバナ 黄色と 多糖類。このうちフェルラ酸は、血小板凝集抑制、抗血栓作用、抗炎症作用、抗酸化作用などの薬理作用を持っています。紅花黄には、抗脳虚血、抗心筋虚血、抗血栓、抗酸化、抗腫瘍などの効果があります。周ら。 THSWDは、脳虚血再灌流傷害後の神経障害機能、脳梗塞容積、脳組織形態を改善し、DRP1、NLRP3、カスパーゼ-1、およびIL-1 βタンパク質のレベルを大幅に低下させることを発見しました。 [233]。これは、脳梗塞に対するTHSWDの治療効果が、パイロトーシスに関連するDRP1/NLRP3経路の阻害に関連していることを示した。さらなる研究により、THSWD グループの IL-1β および IL-18 の含有量が大幅に減少し、NLRP3、カスパーゼ-1、カスパーゼ-1 p10、ASC、TXNIP、および GSDMD のレベルが減少したことが示されました。シグナル伝達経路の検出により、THSWD が HMGB1/RAGE、TLR4/NF-κB、 p38 マップ そして半影にあるJNK。要約すると、THSWD は MCAO/R ラットの炎症反応レベルを低下させ、MCAO/R ラットの NLRP3 インフラマソームの活性化を阻害し、GSMDD を下方制御しました。 THSWD にはピロトーシスを阻害する効果があり、これは HMGB1/TLR4/NF-kB および MAPK シグナル伝達経路の阻害によって影響を受ける可能性があります。 [234].
5.2.2.ナオシンチンカプセル (NXQC)
NXQCの主成分はヤギの葉です。 ディオスピロスカキ トゥンブ(柿の葉)。研究によると、柿の葉エキスにはフラボノイド、有機酸、 クマリン 抗炎症作用、抗酸化作用、その他の化学物質 降圧剤、血中脂質の低下、改善 血管平滑筋 機能と 血行力学的 関数。この薬は心血管疾患や脳血管疾患の治療に広く使用されています。 [235], [236], [237]。既存の研究では、NXQC が効果的に改善できることが示されています。 脳動脈硬化症 脳虚血によって引き起こされる神経損傷を保護します [238]。ミンら。 NXQCは脳虚血後の学習能力の低下を効果的に改善し、脳の活動を増加させることができることを発見しました。 スーパーオキシドジスムターゼ そして 乳酸脱水素酵素、GSHの含有量を増やし、の含有量を減らします。 マロンジアルデヒド。 NXQC はまた、海馬の ASC、NLRP3、およびカスパーゼ-1 タンパク質を下方制御し、IL-18 および IL-1β の含有量を大幅に減少させることができます。彼らはまた、アレチネズミの血小板内皮細胞接着分子-1陽性細胞がNXQCによる介入後に有意に増加したことも発見した。 細胞間結合 きつかったです。 NXQC が細胞の形態を効果的に保護できることが示唆されています。 海馬CA1領域 スナネズミの増殖を抑制し、脳血管機能を保護し、脳虚血再灌流障害を抑制します。 [239].
5.2.3.芙蓉黄湯煎じ薬(BYHWD)とその改良品
BYHWDは有名です 伝統的な中国医学 脳卒中の治療に使用される処方。その処方は清朝の王清仁の『医経正正』に初めて記録された。 [240], [241]。気を養い、血液を活性化する処方の代表的なものです。気を強化し、血液循環を促進し、側副を浚渫する機能があります。科学的根拠に基づいた医学研究で良好な臨床効果が証明されています [242], [243]。私たちの以前の調査では、BYHWD が 配糖体 神経機能不全を改善し、神経細胞の損傷を軽減し、神経細胞のピロトーシスを抑制します。 BYHWD 配糖体は、BYHWD から抽出された有効成分で、主にアストラガロシド IV、ペオニフロリン、 アミグダリン。腎臓を強化し、血液を活性化する主な薬理活性成分であり、脳虚血性神経損傷を治療できます。 [244]。さらに、BYHWD グリコシドが、NLRP3 媒介標準ピロトーシス経路の NLRP3、ASC、プロカスパーゼ-1、カスパーゼ-1、および IL-1β タンパク質の発現を有意に阻害することを観察しました。 [209]。要約すると、BYHWD グリコシドは、CIRI 後のニューロンのパイロトーシスを阻害することで神経保護効果を発揮します。これは、NLRP3 による古典的なパイロトーシス経路の制御と密接に関連しています。さらにロンジーは、 煎じ薬 は、BYHWD がヒルと アキランテス シチュネンシス。急性脳卒中の臨床治療に長年使用されており、その効果は非常に良好です [245]。最近の研究では、龍芝煎じ薬がCIRI後のラットの神経症状を改善し、脳組織の神経細胞の状態を維持し、脳梗塞の容積を減少させる明らかな効果があることが示されています。介在細胞ピロトーシスに関して、龍芝煎じ薬はCIRI後のラットの症状を改善し、保護することができます。 神経細胞の損傷 カスパーゼ-1 および IL-18 タンパク質の発現を下方制御することによる損傷後の損傷 [245].
5.2.4. Yiqi Huoxue 処方箋 (YQHXP)
YQHXP の構成要素 当帰シネンシス基、レンゲ基、 チュアンシオン・リゾマ、蠍座、レオヌリ・ハーブ、アコリ・タタリノウィ・リゾマ、ボルネオラム・シンセティウム、良好な臨床結果を達成しています [246]。動物実験では、YQHXPがラットの神経機能スコアと脳梗塞率を改善し、脳組織のIL-1β、TNF-α、IL-18のレベルとIL-18の相対発現を低下させることが示されています。 P2RX7、脳虚血周囲組織におけるカスパーゼ-1、カスパーゼ-11、およびGSDMD。 YQHXP 介入後、脳組織の炎症性滲出液と浮腫が大幅に減少し、細胞形態と神経空胞変性が改善されました。要約すると、YQHXP はピロトーシスを阻害することにより、急性虚血性脳卒中ラットの脳組織の炎症を軽減する可能性があります。 [247].
5.2.5.沈尚福源煎じ薬 (SNFYD)
SNFYDは腎臓と脳の同時治療理論に基づいた虚血性脳卒中に対する効果的な処方です。その構成は次のとおりです。 黄耆基数 30g, ジオウ 基数プラエパラタ 10g、コルニフルクトゥス 10g、ヤマノイモ科の根茎 15g、イワベンケイ、イワベンケイ、根茎および根茎 20g, ムータン皮質 10g、アンジェリカエ・シネンシス・ラディックス 10g、ボタン、Radix Rubra 10g, フェレティマ 10g [248]。血液を活性化して側副血行路を浚渫する、気を元気にする薬、エッセンスを補充する薬、腎臓を元気にする薬が多数配合されています。研究では、OGD モデリング後、PC12 が大幅に損傷し、パイロトーシス経路のさまざまなサイトカインが大幅に増加したことが示されています。 [249]。 SNFYD含有血清とINF39が損傷部位に介入した後 PC12セル、PC12細胞の形態と活性が改善され、それに応じてNLRP3/カスパーゼ-1ピロトーシス経路の活性化が弱まりました。これは、この経路の阻害が細胞状態の改善に関連しており、SNFYD と INF39 の両方がこの経路の活性化を阻害できることを示しています。 SNFYDは、NLRP3/カスパーゼ-1および下流のパイロトーシス経路を阻害することにより神経細胞の炎症性アポトーシスを軽減し、それによって神経組織を保護し、脳梗塞の治療という目的を達成する可能性があることが示唆されています。 [250].
5.2.6.その他の抜粋
さらに、最近の研究では、ヘパリンなどの抗血栓薬がピロトーシスを抑制することで肺損傷を改善できることが示されています。例えば、ヤンら。ヘパリンが hMGB1 LPS 誘導性のカスパーゼ 11 活性化をブロックすることで肺内皮細胞のアポトーシスを阻害することを発見しました。これは、敗血症誘導性の肺損傷を治療する潜在的な方法となる可能性があります。 [251]。プラタパンら。柔らかいココナッツウォーターには抗酸化作用と抗血栓作用があることがわかりました。 実験的心筋梗塞 モデル [252]一方、若いココナッツジュースは、に関連する病状の一部を大幅に軽減することができます。 アルツハイマー病 [253]。ココナッツウォーターはパイロトーシスの潜在的な阻害剤である可能性が示されており、ISにおけるパイロトーシスの制御におけるココナッツウォーターの役割は将来研究される可能性があります。
L-アルギニンは、体内に存在する有機化合物です。 プロタミン 大量に [254]。それは合成の前駆体です。 一酸化窒素 (NO) は血管の無傷の内皮を保護し、 血管拡張剤 および内因性の抗アテローム性動脈硬化分子 [254], [255]。動物実験でわかったことは、 L-アルギニンは複合体を持っている可能性があります 抗凝固作用、抗凝固作用、および 線維素溶解 効果 [255], [256]。高血圧ラットでは、Cylwik et al.あれを見つけた L-アルギニンはラットの血圧を下げる可能性があり、長期治療によりユーグロブリン血栓溶解時間と出血時間が短縮され、コラーゲン誘発性の血小板凝集が抑制されました。示唆されているのは、 L-アルギニンは高血圧ラットモデルにおいて抗血栓作用を果たします。 静脈血栓症 複雑な方法で [256]。最近の研究では、 L-アルギニンはパイロトーシスの潜在的な阻害剤である可能性がある: Tanuseputero et al.と提案した L-アルギニンはNLRP3インフラマソームを部分的に阻害して敗血症誘発性を軽減する可能性がある 急性腎障害 マウスで [257]。血管拡張作用、抗血栓作用、および NLRP3 インフラマソーム活性の阻害を考慮すると、 L-アルギニン、と示唆されています。 L-アルギニンは、虚血領域の血流を増加させ、ISのパイロトーシスを抑制する可能性があります。
パイロトーシスを制御する天然物の概要を以下に示します。 表2.
表2。パイロトーシスを制御する天然化合物の概要。
| 天然物 | モデル/疾患 | 種 | 効果 | 参照 |
|---|---|---|---|---|
| ガストロディン | 中大脳動脈閉塞/再灌流 (MCAO/R)、酸素欠乏/再酸素化 (OGD/R) | ドブネズミとドブネズミの縁皮質ニューロン | NLRP3、炎症因子 (IL-1β、IL-18) および切断型カスパーゼ 1 を下方制御することによりパイロトーシスを抑制します。 lncRNA NEAT1/miR-22–3p 軸および lncRNA NEAT1/miR-22–3p 軸を制御します | [204] |
| タンシノーネIIA | OGD/R | BV2細胞(ハツカネズミ由来のミクログリア) | NLRP3 炎症シグナル伝達経路における IL-1β および IL-18 の発現を低減します。 | [205] |
| レオヌリン | マカオ | ドブネズミ | NLRP3の発現を低下させます | [206] |
| ペオニフロリン | マカオ | ドブネズミ | NLRP3、カスパーゼ-1、IL-1βレベルを阻害 | [209] |
| ブチルフタリド | マカオ | ドブネズミ | NLRP3 インフラマソームシグナル伝達経路を調節します | [210] |
| ブレビスカピン | MCAO/R | ドブネズミ | CCI ラットの海馬における NLRP3 炎症細胞の活性化を阻害し、カスパーゼ 1、IL-6 および IL-1β タンパク質の発現を下方制御し、カスパーゼ 3 タンパク質の活性化を阻害します。 | [211] |
| レスベラトロール | MCAO/R | ドブネズミ | ミクログリア NLRP3 インフラマソーム、カスパーゼ-1、ZO-1 を制御します | [212] |
| サルビアノール酸 | MCAO/R および OGD/R | ドブネズミとドブネズミのミクログリア | NLRP3 インフラマソームの活性化と NLRP3 インフラマソーム/発熱関連タンパク質の発現を阻害します。 | [213] |
| ヒスピドゥリン | マカオ | ドブネズミ | AMPK/GSK3βシグナル伝達経路を調節することにより、NLRP3媒介ピロトーシスを阻害します | [214] |
| アストラガロシド IV | MCAO/R | ドブネズミ | NLRP3 インフラマソームの活性化を阻害します。脳組織におけるNLRP3、カスパーゼ-1、プロIL-1β、IL-1β、プロIL-18およびIL-18のタンパク質レベルを低下させます。リン酸化NF-κBタンパク質の発現を阻害します。 | [223] |
| オタネニンジンサポニン | MCAO/R | ドブネズミ | PINK1/パーキン経路を調節します。 NLRP3 インフラマソームの活性化を阻害します | [232] |
| 桃紅四烏煎じ薬 | マカオ | ドブネズミ | DRP1、NLRP3、カスパーゼ-1、IL-1 βタンパク質のレベルを低下させます。 | [233] |
| 桃紅四烏煎じ薬 | MCAO/R | ドブネズミ | HMGB1/TLR4/NF-kBおよびMAPKシグナル伝達経路を阻害します | [234] |
| ナオシンチンカプセル | MCAO/R | メリオネス・ウングキュラトゥス | ASC、NLRP3、およびカスパーゼ-1 タンパク質を下方制御します。 IL-18およびIL-1βの含有量を減少させます。 | [239] |
| 芙楊黄粉煎じ薬 | MCAO/R | ドブネズミ | NLRP3 媒介標準ピロトーシス経路の NLRP3、ASC、プロカスパーゼ-1、カスパーゼ-1、および IL-1β タンパク質の発現を阻害します。 | [244] |
| 龍芝煎じ薬 | MCAO/R | ドブネズミ | カスパーゼ-1 および IL-18 タンパク質の発現を下方制御します | [245] |
| Yiqi Huoxueの処方箋 | マカオ | ドブネズミ | IL-1β、TNF-α、IL-18のレベルと、P2RX7、カスパーゼ-1、カスパーゼ-11、GSMDDの相対発現を低下させます。 | [247] |
| 沈尚夫源煎じ薬 | OGD | ドブネズミ由来の PC12 細胞 | NLRP3/カスパーゼ-1を阻害します | [250] |
6. 展望
私たちの研究チームは、IS後の血管神経単位におけるさまざまな細胞群(ニューロン、ミクログリア、アストロサイトなど)のプログラムされた細胞死様式を長年研究してきました。他の研究報告と組み合わせると、アポトーシス、フェロトーシス、オートファジーなどのさまざまな細胞死モードがIS後に発生し、アポトーシスとパイロトーシスの形態には一定の類似性があることがわかりました。 神経炎症 IS 後は脳損傷につながる最も重要な病理学的プロセスであり、パイロトーシスは神経炎症の重要な部分を占めます。これは、パイロトーシスが脳虚血後の神経炎症関連プログラム細胞死において異常に重要であることを示唆している。パイロトーシスによって引き起こされる関連シグナル伝達経路は、他のプログラム細胞死経路とも交差しており、パイロトーシスを標的とした薬剤開発がISの重要な治療法であることが示唆されています。
現在の研究には、まだ改善の余地があります: (1) 現在の研究の動物モデルにおける正規化と統一の問題により、動物のさまざまな細胞におけるパイロトーシスに関する結論は、 神経系 矛盾しています。したがって、将来のパイロトーシスの研究には、抗パイロトーシス薬の実際の効果を決定することもできる、虚血性脳卒中の動物モデルにおけるより標準化され統一された安定したモデルが必要です。 (2) 今後の研究では、天然化合物の長期的な脳保護効果と、さまざまなげっ歯類および大型哺乳類の脳卒中モデルにおけるパイロトーシスを制御するメカニズムを検証できる可能性があります。今後はさらに注意を払う必要がある 薬物動態, 薬力学、および現在パイロトーシスを調節している天然化合物の毒性学的特性。 (3) さらに、脳虚血性脳卒中におけるパイロトーシスの制御における天然化合物の組み合わせの相乗効果と、神経炎症を抑制する神経保護効果についても、将来的に調査する必要がある。 (4) すべての結果が良好であれば、次のステップは次のステップです。 臨床試験 潜在的な ファイトケミカル 脳虚血/脳卒中に対する神経保護効果を調査するためです(たとえば、チームは現在、脳小血管疾患の治療におけるNaotai Fangの臨床試験を実施中です: ChiCTR1900024524)。
7. まとめ
炎症促進性のプログラム細胞死であるパイロトーシスは、主にカスパーゼ-1によって媒介される古典的なパイロトーシス経路を介して細胞死と神経炎症を誘導することにより、虚血性脳卒中の病理学的プロセスにおいて重要な役割を果たしています。非標準的なパイロトーシス経路と IS との関係はまだ解明されていません。パイロトーシスの他のメカニズムについては、今後さらなる研究が必要です。現在、研究により次のことが確認されています。 薬物標的化 パイロトーシス経路の主要なタンパク質は虚血性脳損傷をある程度緩和することができますが、その研究は主に細胞実験と動物実験に限定されており、研究は不足しています。 臨床研究 証拠。したがって、ISにおけるピロトーシスの制御機構を探求するさらなる臨床研究により、虚血性脳卒中を予防および治療するための新しい治療戦略と理論的基礎が提供されることが期待されている。天然化合物によるパイロトーシスの制御に関して、我々の一般化を通じて、天然植物化合物は単一成分だけでなくパイロトーシスを制御することによって脳虚血を制御できることが分かる。多成分の天然植物化合物は、パイロトーシスによって引き起こされる炎症カスケードの調節において潜在的な相乗効果を示すこともわかります。このことは、将来、さまざまなシグナル伝達経路によって引き起こされるピロトーシス媒介神経炎症に対するさまざまな天然化合物の制御効果を研究することへの動機付けとなります。
CRediT著者寄稿文
ヤン・カイリン: 概念化、方法論、形式的分析、調査、執筆 - 原案。 リウティン・ゼン: 概念化、方法論、ソフトウェア、形式分析、調査、執筆 - オリジナルの草案。 ジンソン・ゼン: 方法論、形式的な分析、調査。 ティンティン・バオ: 方法論、形式的分析、調査、執筆 - 原案。 シャオユアン: 方法論、形式的な分析、調査。 シャンシャン・ワン: 方法論、形式的な分析、調査。 王翔: 方法論、形式的な分析、調査。 ハオ・シュウ: 方法論、形式的な分析、調査。 ジンウェン・ゲー: 概念化、方法論、形式分析、調査、執筆 - レビューと編集。
利益相反に関する声明
著者らは、この論文で報告されている研究に影響を与えた可能性がある既知の競合する経済的利益や個人的関係を持っていないことを宣言します。
謝辞
この作品は、 中国国家自然科学財団 (81774174)、 中国の国家重点研究開発プロジェクト (いいえ。 2018YFC1704904), 湖南省国立自然科学財団、 中国 (2020JJ5424 そして 2020JJ5442), 湖南中医薬大学「ダブルファーストクラス」中国伝統医学と西洋医学の統合の規律オープンファンドプロジェクト (2020ZXYJH08 そして 2020ZXYJH09), 湖南省教育局青少年基金プロジェクト (21B0386).
データと資料の入手可能性
この研究の結果を裏付けるために使用されたデータは、記事内に含まれています。
参考文献
- [1]
心血管疾患、炎症、脳小血管疾患Cardiovasc Res.、117 (13) (2021)、pp. 2575-2588、 10.1093/cvr/cvab284
- [2]
脳卒中に対する運動効果の臨床的証拠上級経験値医学。 Biol.、1000 (2017)、131-151 ページ、 10.1007/978-981-10-4304-8_9
- [3]
世界脳卒中機関 (WSO): 世界脳卒中ファクトシート 2022内部。 J. Stroke、17 (1) (2022)、pp. 18-29、 10.1177/17474930211065917
- [4]
小児期の脳卒中ナット。 Rev.Dis. Prim.、8 (1) (2022)、p. 12、 10.1038/s41572-022-00337-x
- [5]
虚血および再灌流傷害における現在の機構概念Cell PhysiolBiochem、46 (4) (2018)、pp. 1650-1667、 10.1159/000489241
- [6]
脳卒中の疫学と一次予防コンティン。 (ミンネ)、26 (2) (2020)、pp. 260-267、 10.1212/CON.0000000000000831
- [7]
若年成人における虚血性脳卒中の疫学、病因学、および管理Lancet Neurol.、17 (9) (2018)、pp. 790-801、 10.1016/S1474-4422(18)30233-3
- [8]
中国の脳卒中研究の協力。中国の脳卒中:疫学、予防、管理における進歩と課題Lancet Neurol.、18 (4) (2019)、pp. 394-405、 10.1016/S1474-4422(18)30500-3
- [9]
虚血/再灌流関連傷害におけるフェロトーシスの病理学的役割ズール。 Res、41 (3) (2020)、pp. 220-230、 10.24272/j.issn.2095-8137.2020.042
- [10]
虚血および再灌流傷害における現在のメカニズムの概念Cell PhysiolBiochem、46 (4) (2018)、pp. 1650-1667、 10.1159/000489241
- [11]
フェロトーシスを標的とする:虚血再灌流傷害の病理学的メカニズムと治療オキシド。 Med Cell Longev.、2021 (2021)、記事 1587922、 10.1155/2021/1587922
- [12]
脳卒中の発症における新たなエピジェネティックな役割: ノンコーディング RNA からエキソソームのノンコーディング RNA へバイオメッド。薬学、140 (2021)、記事 111753、 10.1016/j.biopha.2021.111753
- [13]
脳梗塞/虚血再灌流傷害における神経炎症および免疫調節におけるノンコーディングRNAの研究進歩の系統的レビューフロント。免疫学、13 (2022)、記事 930171、 10.3389/fimmu.2022.930171
- [14]
虚血性脳卒中における酸化ストレスを制御するフェロトーシスの機構と天然薬理活性成分の制御機構バイオメッド。薬学、154 (2022)、記事 113611、 10.1016/j.biopha.2022.113611
- [15]
炎症と脳卒中のリスク: 予防の新たな目標脳卒中、52 (8) (2021)、pp. 2697-2706、 10.1161/ストロークアハ.121.034388
- [16]
脳卒中研究のギャップを埋める: 炎症と免疫に焦点を当てる脳の行動。 Immun.、91 (2021)、pp. 649-667、 10.1016/j.bbi.2020.09.025
- [17]
パイロトーシス:ガスデルミン媒介のプログラムされた壊死性細胞死Trends Biochem Sci.、42 (4) (2017)、pp. 245-254、 10.1016/j.tibs.2016.10.004
- [18]
パイロトーシス:がんの新境地バイオメッド。薬学、121 (2020)、記事 109595、 10.1016/j.biopha.2019.109595
- [19]
神経変性疾患の治療におけるNLRP3インフラマソームの関与バイオメッド。薬学、138 (2021)、記事 111428、 10.1016/j.biopha.2021.111428
- [20]
卵巣がんにおける非アポトーシス細胞死:治療、耐性および予後バイオメッド。薬学、150 (2022)、記事 112929、 10.1016/j.biopha.2022.112929
- [21]
てんかん重積状態における細胞死経路のメカニズムと関連治療薬バイオメッド。薬学、149 (2022)、記事 112875、 10.1016/j.biopha.2022.112875
- [22]
脳卒中におけるピロトーシス - 疾患メカニズムと治療戦略に関する新たな洞察J. PhysiolBiochem、77 (4) (2021)、pp. 511-529、 10.1007/s13105-021-00817-w
- [23]
ピロトーシスを標的にして虚血性脳卒中損傷を制御する: 分子機構と前臨床証拠Brain Res Bull.、165 (2020)、pp. 146-160、 10.1016/j.brainresbull.2020.10.009
- [24]
脳虚血再灌流のラットモデルにおけるピロトーシスおよびネクロトーシスに対するアストラガロシドIVとオタネニンジンサポニンの組み合わせの効果経験値それで。 Med、22 (4) (2021)、p. 1123、 10.3892/etm.2021.10557
- [25]
ラットの脳虚血再灌流傷害後のパイロトーシスに対するBuyangHuanwu煎じ薬に含まれる配糖体の神経保護効果J. Ethnopharmacol.、242 (2019)、記事 112051、 10.1016/j.jep.2019.112051
- [26]
唐彪、唐文静、戴子偉、沙徐、鄧長清。オタネニンジンサポニンは、SH-SY5Y 細胞の低酸素性低血糖/再酸素化および再血糖誘発性のパイロトーシスを阻害します。中国病態生理学ジャーナル、2020、36 (07):1178–1184.(中国語)。
- [27]
細胞パイロトーシスのメカニズムに基づいて、低血糖および低酸素状態にある PC12 細胞に対する神納富源煎じ薬の保護効果を調査する [D]湖南大学伝統。顎。医学。 (2019)、 10.27138/d.cnki.ghuzc.2019.000152(中国語で)
- [28]
Chaihu-Longgu-Muli煎じ薬は、海馬ニューロンのパイロトーシスを改善することにより、ラットの抗てんかん効果を発揮しますJ. Ethnopharmacol.、270 (2021)、記事 113794、 10.1016/j.jep.2021.113794
- [29]
細胞死の分子機構: 2018 年細胞死命名委員会の勧告Cell Death Different.、25 (2018)、pp. 486-541
- [30]
健康と病気におけるアポトーシスの制御: BCL-2 ファミリータンパク質のバランス作用ナット。モル牧師。 Cell Biol.、20 (2019)、pp. 175-193
- [31]
パイロトーシス:メカニズムと病気信号伝達。 Target Ther.、6 (1) (2021)、p. 128、 10.1038/s41392-021-00507-5
- [32]
パイロトーシスは非選択的ガスダーミン D 細孔によって引き起こされ、その形態は MLKL チャネル媒介ネクロトーシスとは異なります。Cell Res、26 (2016)、pp. 1007-1020
- [33]
調節された細胞死の分子機構Cell Res、29 (5) (2019)、pp. 347-364
- [34]
NLRC4 インフラマソームを介した III 型分泌装置の自然免疫検出手順国立アカド。サイエンス、107 (7) (2010)、pp. 3076-3080
- [35]
カスパーゼ-1 ダイジェトームは、感染および敗血症性ショック時の標的として解糖経路を特定しますJ.Biol. Chem.、282 (50) (2007)、pp. 36321-36329
- [36]
化学療法薬はガスダーミンのカスパーゼ 3 切断を通じてパイロトーシスを誘発しますネイチャー、547 (7661) (2017)、99-103 ページ
- [37]
BCL-2タンパク質ファミリーによるアポトーシスの制御: 生理学と治療への影響Nature Rev. Mol. Cell Biol.、15 (1) (2014)、pp. 49-63
- [38]
アポトーシスの分子機構とがんの発症と治療における役割アジアンパックJ. Cancer Prev.、16 (6) (2015)、pp. 2129-2144
- [39]
治療用小分子は、外因性および内因性細胞死経路の調節解除を伴う癌におけるアポトーシスタンパク質の阻害剤を標的とするクリン。 Cancer Res.、23 (6) (2017)、pp. 1379-1387
- [40]
ネクロプトーシスN.Engl. J. Med.、370 (5) (2014)、455-465 ページ
- [41]
シュードキナーゼMLKLは分子スイッチ機構を介してネクロトーシスを媒介する免疫、39 (3) (2013)、pp. 443-453
- [42]
代替細胞死経路と細胞代謝内部。 J. Cell Biol.、2013 (2013)、記事 463637
- [43]
プログラムされた細胞死の間に原形質膜が変化するCell Res、28 (1) (2018)、pp. 9-21
- [44]
細胞死における鉄と活性酸素種の役割ナット。化学。 Biol.、10 (2014)、pp. 9-17、 10.1038/nchembio.1416
- [45]
Bogdan AR、宮沢 M.、橋本 K.、辻 Y. 鉄ホメオスタシスの調節因子: 代謝、細胞死、および疾患における新たな役割。
- [46]
フェロトーシスと自己免疫疾患フロント免疫、13 (2022)、記事 916664、 10.3389/fimmu.2022.916664PMID: 35720308; PMCID: PMC9203688。
- [47]
血球関連疾患のフェロトーシスを標的とするJ. 薬物ターゲット、30 (3) (2022)、pp. 244-258、 10.1080/1061186X.2021.1971237
- [48]
心血管疾患におけるパイロトーシスの役割Int Immunopharmacol.、67 (2019)、pp. 311-318、 10.1016/j.intimp.2018.12.028
- [49]
NLRP3/ASC媒介肺胞マクロファージピロトーシスは、心肺バイパスによって誘発された急性肺損傷におけるHMGB1分泌を亢進する研究室調査します。 ; J. Tech. Methods Pathol.、98 (8) (2018)、pp. 1052-1064、 10.1038/s41374-018-0073-0
- [50](a)
NLR インフラマソームの構築とシグナル伝達の構造メカニズムカー。上。微生物免疫学、397 (2016)、pp. 23-4210
(b)
インフラマソーム集合の構造機構FEBS J.、282 (3) (2015)、pp. 435-444 - [51](a)
インフラマソーム複合体: 新たなメカニズムとエフェクター機能セル、165 (4) (2016)、pp. 792-8007
(b)
多機能炎症メディエーターとしてのカスパーゼ-1: 非サイトカイン成熟の役割J.ロイコック。 Biol.、100 (5) (2016)、pp. 961-967 - [52]
自己免疫疾患におけるNLRP3インフラマソームの役割バイオメッド。薬学、130 (2020)、記事 110542、 10.1016/j.biopha.2020.110542
- [53]
インフラマソーム活性化の制御イムノール。 Rev.、265 (1) (2015)、6-21 ページ、 10.1111/imr.12296
- [54]
ユビキチンシステムによるインフラマソームの制御FEBS J.、287 (1) (2020)、pp. 11-26、 10.1111/2月15118
- [55]
インフラマソームシグナル伝達の分子機構J.ロイコック。 Biol.、103 (2) (2018)、pp. 233-257、 10.1189/jlb.3MR0617-250R
- [56]
インフラマソームの操作: 肺がんの免疫療法への有望なアプローチInt Rev. Immunol.、40 (3) (2021)、pp. 171-182、 10.1080/08830185.2021.1876044
- [57]
パネキシン-1はアポトーシス中のATP放出に必要ですが、インフラマソームの活性化には必要ではありませんJ. Immunol.、186 (11) (2011)、pp. 6553-6561、 10.4049/ジムノール.1100478
- [58]
NLRP3 インフラマソーム: 活性化と制御のメカニズムの概要Int J.Mol.科学、20 (13) (2019)、p. 3328、 10.3390/ijms20133328
- [59]
NLRP3 インフラマソームと婦人科疾患および産科合併症におけるその重要な役割フロント免疫、11 (2021)、記事 555826、 10.3389/fimmu.2020.555826
- [60]
出血性脳卒中の病態生理学における NLRP3 インフラマソーム: 総説カー。 Neuropharmacol.、17 (7) (2019)、pp. 582-589、 10.2174/1570159X17666181227170053
- [61]
カスパーゼ-1は、カスパーゼのリクルートと基質切断を媒介する2つの異なる界面を介して全長ガスダーミンDに関与するe5免疫、53 (1) (2020)、pp. 106-114、 10.1016/j.immuni.2020.06.007
- [62]
カスパーゼ切断モチーフ抗体の発見により、非標準的なインフラマソーム機能に関する洞察が明らかに手順国立アカド。科学。米国、118 (12) (2021)、記事 e2018024118、 10.1073/pnas.2018024118
- [63]
非標準的なカスパーゼ-4/5/11 インフラマソームの機能と制御J. Immunol.、204 (12) (2020)、pp. 3063-3069、 10.4049/jimmunol.2000373PMID: 32513874
- [64]
カスパーゼ切断モチーフ抗体の発見により、非標準的なインフラマソーム機能に関する洞察が明らかに手順国立アカド。科学。米国、118 (12) (2021)、記事 e2018024118、 10.1073/pnas.2018024118
- [65]
非標準的なインフラマソーム媒介ピロトーシスのメカニズムと結果J.Mol. Biol.、434 (4) (2022)、記事 167245、 10.1016/j.jmb.2021.167245
- [66]
カスパーゼ 11 は、パイロトーシスと内毒素ショックを媒介するためにパネキシン 1 チャネルとプリン作動性 P2X7 細孔を必要とします免疫、43 (5) (2015)、pp. 923-932、 10.1016/j.immuni.2015.10.009
- [67]
サクシネーションはガスダーミン D を不活性化し、パイロトーシスをブロックしますサイエンス、369 (6511) (2020)、pp. 1633-1637、 10.1126/science.abb9818
- [68]
パイロトーシスにおけるガスダーミン Dアクタファーム。罪。 B.、11 (9) (2021)、pp. 2768-2782、 10.1016/j.apsb.2021.02.006
- [69]
カスパーゼ-11は非標準的なインフラマソームシグナル伝達のためにガスダーミンDを切断するネイチャー、526 (7575) (2015)、666-671 ページ、 10.1038/自然15541
- [70]
インフラマソーム活性化ガスダーミン D は膜孔を形成することでピロトーシスを引き起こすネイチャー、535 (7610) (2016)、153-158 ページ、 10.1038/自然18629
- [71]
パイロトーシス: ガスダーミン媒介のプログラム壊死性細胞死Trends Biochem Sci.、42 (4) (2017)、pp. 245-254、 10.1016/j.tibs.2016.10.004
- [72]
GSDMD 膜細孔形成 - パイロプトーシス細胞死のメカニズムを構成するEMBO J.、35 (16) (2016)、1766-1778 ページ
- [73]
ピロトーシスとガスダーミンファミリーの機能の根底にある分子機構に関する最近の洞察トレンド免疫学、38 (2017)、pp. 261-271
- [74]
CD T 細胞死を引き起こす優勢なカスパーゼ 1 媒介ピロトーシス: 非局所空間数学モデル Jブル。数学。 Biol.、80 (3) (2018)、pp. 540-582、 10.1007/s11538-017-0389-8
- [75]
長いノンコーディング RNA とパイロトーシスクリン。チム。 Acta、504 (2020)、201-208 ページ、 10.1016/j.cca.2019.11.035
- [76]
糖尿病性心筋症の発症機序におけるノンコーディングRNAの関与J.Cell Mol. Med、23 (9) (2019)、pp. 5859-5867、 10.1111/jcmm.14510
- [77]
長い非コードRNAは、競合する内因性RNAとして機能することで筋肉の分化を制御するセル、147 (2) (2011)、pp. 358-369、 10.1016/j.cell.2011.09.028
- [78]
LncRNA H19は網膜虚血/再灌流傷害におけるミクログリアのピロトーシスとニューロン死を引き起こすCell Death Differ.、27 (1) (2020)、pp. 176-191、 10.1038/s41418-019-0351-4
- [79]
カスパーゼ-8は急性緑内障におけるNLRP1/NLRP3インフラマソーム活性化とIL-1β産生を促進する手順国立アカド。科学。 USA、111 (30) (2014)、pp. 11181-11186、 10.1073/pnas.1402819111
- [80]
H19 lincRNA は、増殖と Igf1r を抑制する miR-675 の発生リザーバーです。ナット。 Cell Biol.、14 (7) (2012)、pp. 659-665、 10.1038/ncb2521
- [81]
H19 は腹部大動脈瘤の発生と進行を誘発する循環、138 (15) (2018)、pp. 1551-1568、 10.1161/循環AHA.117.032184
- [82]
低酸素症および虚血誘発性神経細胞死におけるRip2/カスパーゼ-1経路の基本的な役割手順国立アカド。科学。 USA、100 (26) (2003)、pp. 16012-16017、 10.1073/pnas.2534856100
- [83]
虚血性脳卒中における神経細胞死のメカニズムとその治療的意義Med Res Rev.、42 (1) (2022)、pp. 259-305、 10.1002/med.21817
- [84]
長いノンコーディングRNA MEG3はmiR-485/AIM2軸を標的とすることでピロトーシスを増加させ、脳虚血再灌流障害を促進する経験値Neurol.、325 (2020)、記事 113139、 10.1016/j.expneurol.2019.113139
- [85]
LncRNA-Fendrr は、HERC2 を介して NLRC4 タンパク質のユビキチン化と分解を防ぎ、ミクログリアのパイロトーシスを制御しますモル。 Med、27 (1) (2021)、p. 39、 10.1186/s10020-021-00299-y
- [86]
ガストロジンはlncRNA NEAT1/miR-22-3p軸を調節してピロトーシスを抑制し、脳虚血/再灌流障害を軽減するNeurochem Res、46 (7) (2021)、pp. 1747-1758、 10.1007/s11064-021-03285-2
- [87]
HBO は lncRNA-H19/miR-423-5p/NLRP3 軸を介して神経幹細胞パイロトーシスを軽減し、酸素グルコース欠乏後の神経新生を改善しますオキシド。 Med Cell Longev.、2022 (2022)、記事 9030771、 10.1155/2022/9030771
- [88]
miR-21 は NLRP3 インフラマソームの活性化を促進し、パイロトーシスと内毒素ショックを媒介しますCell Death Dis.、10 (6) (2019)、p. 461、 10.1038/s41419-019-1713-z
- [89]
A20によるNLRP3インフラマソームの負の制御により関節炎を防ぐネイチャー、512 (7512) (2014)、69-73 ページ、 10.1038/自然13322
- [90]
炎症性カスパーゼによるGSDMDの切断がパイロトーシス性細胞死を決定するネイチャー、526 (7575) (2015)、660-665 ページ、 10.1038/自然15514
- [91]
LncRNA KCNQ1OT1 は糖尿病性心筋症におけるピロトーシスを媒介する細胞生理学。 Biochem、50 (4) (2018)、pp. 1230-1244、 10.1159/000494576
- [92]
細胞死の分子機構: 2018 年細胞死命名委員会の勧告細胞死の差異、25 (3) (2018)、pp. 486-541、 10.1038/s41418-017-0012-4
- [93]
オートファジーの薬理学的調節: 治療の可能性と持続する障害。Nat Rev Drug Discov。 2017;16(7):487–511。
- [94]
オートファジー: 細胞と組織の再生セル、147 (4) (2011)、pp. 728-741
- [95]
AIM2 および NLRP3 インフラマソームは、ASC を介してアポトーシスとパイロトーシスの両方の死経路を活性化します細胞死差、20 (9) (2013)、pp. 1149-1160、 10.1038/cdd.2013.37
- [96]
NLRP3 欠失は高酸素誘発性の急性肺損傷から保護します午前。 JPhysiol, 305 (1 Pt.1) (2013), pp. C182-C189, 10.1152/ajpcell.00086.2013
- [97]
NLRP3 タンパク質発現およびイースパーゼ 1 媒介インターロイキン 1 β 分泌の制御におけるリポ多糖/アデノシン三リン酸媒介シグナル伝達インフラム。 Res、62 (1) (2013)、pp. 89-96、 10.1007/s00011-012.0555.2
- [98]
メラトニンはマウス脂肪組織におけるNF-κB/GSDMDシグナルを阻害することで、インフラトーム誘発性ピロトーシスを軽減するJPinealRes、63 (1) (2017)、p. 7、 10.1111/jpi.12414
- [99]
急性虚血性脳卒中における炎症とそのメディエーターの役割J. Cardiovasc Transl. Res、6 (5) (2013)、pp. 834-851、 10.1007/s12265-013-9508-6
- [100]
虚血性脳卒中における神経炎症メカニズム: 心塞栓性脳卒中、背景、および治療法に焦点を当てるInt J.Mol.科学、21 (18) (2020)、p. 6454、 10.3390/ijms21186454
- [101]
虚血性脳卒中における脂質メディエーターと無菌炎症Int Immunol.、32 (11) (2020)、pp. 719-725、 10.1093/intimm/dxaa027
- [102]
グリア細胞:虚血性脳卒中における免疫応答の役割Front Immunol.、11 (2020)、p. 294、 10.3389/fimmu.2020.00294
- [103]
MCC950によるNLRP3インフラマソーム活性化の阻害は、db/dbマウスの糖尿病性脳症を改善する分子、23 (3) (2018)pii:E522
- [104]
miRNA-223-3p は NLRP3 を調節してアポトーシスを促進し、hep3B 細胞の増殖を阻害します経験値それで。メッド、15 (3) (2018)2 429-2 435
- [105]
NLRP3 インフラマソームは気管支肺異形成の発症に決定的に関与していますナット。 Commun.、6 (2015)、p. 8977、 10.1038/ncomms9977
- [106]
シンバスタチンは NLRP3 インフラマソーム活性化を阻害し、KLF2 媒介メカニズムにより高酸素誘発性気管支肺異形成症における肺損傷を改善しますオキシド。 Med Cell Longev.、2022 (2022)、記事 8336070、 10.1155/2022/8336070
- [107]
核 DNA 損傷によって引き起こされる NLRP3 インフラマソーム活性化は、ヒト角化細胞における UVB 誘発炎症反応を促進する生化学生物物理学。 Res Commun.、477 (3) (2016)、pp. 329-335、 10.1016/j.bbrc.2016.06.106
- [108]
NLRP3/ASC媒介肺胞マクロファージピロトーシスは、心肺バイパスによって誘発された急性肺損傷におけるHMGB1分泌を亢進するLab Invest、98 (8) (2018)、pp. 1052-1064、 10.1038/s41374-018-0073-0
- [109]
T細胞のインフラマソームJ.Mol. Biol.、434 (4) (2022)、記事 167275、 10.1016/j.jmb.2021.167275
- [110]
NLRP3 インフラマソームは膀胱出口閉塞によって引き起こされる炎症を仲介するJ. Urol.、195 (5) (2016)、pp. 1598-1605、 10.1016/j.juro.2015.12.068
- [111]
神経系疾患のトランスレーショナル治療における NLRP3 インフラマソームの標的化: 最新情報Front Pharm.、12 (2021)、記事 707696、 10.3389/fphar.2021.707696
- [112]
遠隔虚血コンディショニングは炎症を軽減しますか?自然免疫とサイトカイン反応に焦点を当てるBasic Res Cardiol.、116 (1) (2021)、p. 12、 10.1007/s00395-021-00852-0
- [113]
HDAC3阻害剤RGFP966は、AIM2インフラマソームを下方制御することにより虚血性脳損傷を改善したFASEB J.、34 (1) (2020)、pp. 648-662、 10.1096/fj.201900394RRR
- [114]
脳梗塞におけるミクログリアの代謝障害と神経炎症J.Pharm. Sci.、145 (1) (2021)、pp. 130-139、 10.1016/j.jphs.2020.11.007
- [115]
ミトコンドリア機能不全は脳虚血/再灌流傷害中にNLRP3インフラマソーム活性化を誘導するJ. Neuroinflamm.、15 (1) (2018)、p. 242、 10.1186/s12974-018-1282-6
- [116]
脳虚血/再灌流損傷におけるミトコンドリア複合体I損傷のメカニズム。仮説モル。 Cell Neurosci.、100 (2019)、記事 103408、 10.1016/j.mcn.2019.103408
- [117]
酸化ストレス誘発性パイロトーシスはMG63細胞の骨形成機能不全を引き起こすJ.Mol. Histol.、51 (3) (2020)、pp. 221-232、 10.1007/s10735-020-09874-9
- [118]
内皮機能不全における NLRP3 インフラマソームCell Death Dis.、11 (9) (2020)、p. 776、 10.1038/s41419-020-02985-x
- [119]
亜鉛と活性酸素種の間の相乗相互作用がラットの虚血性脳損傷を増幅する脳卒中、49 (9) (2018)、pp. 2200-2210、 10.1161/ストロークアハ.118.021179
- [120]
H9C2心筋細胞におけるROS依存性NLRP3インフラマソーム媒介ピロトーシスの活性化により、高血糖および低酸素/再酸素化誘発損傷を悪化させるJ. Diabetes Res、2019 (2019)、記事 8151836、 10.1155/2019/8151836
- [121]
GSDMD酸化を誘導することによりマクロファージのピロトーシスを促進するJ.Mol. Cell Biol.、11 (12) (2019)、pp. 1069-1082、 10.1093/jmcb/mjz020
- [122]
虚血再灌流傷害におけるフリーラジカル損傷:血行再建療法後の急性虚血性脳卒中における障害オキシド。 Med Cell Longev.、2018 (2018)、記事 3804979、 10.1155/2018/3804979
- [123]
酵母Vps1pおよびDnm1pと相同な新規哺乳類ダイナミン関連タンパク質の同定と細胞内局在J. Biochem、122 (3) (1997)、pp. 525-530、 10.1093/oxfordjournals.jbchem.a021784
- [124]
ネクロスタチン-1はDrp1発現の減少を通じてラット尿細管細胞株NRK-52Eにおける虚血傷害誘発細胞死を軽減するInt J.Mol. Sci.、14 (12) (2013)、pp. 24742-24754、 10.3390/ijms141224742
- [125]
ヒト腎尿細管上皮細胞の低酸素症および再酸素化誘発性パイロトーシスのメカニズムにおけるダイナミン-1 の役割 [D]第三陸軍医科大学 (2016)
- [126]
ミトコンドリア機能不全は脳虚血/再灌流傷害中にNLRP3インフラマソーム活性化を誘導するJ. Neuroinflamm.、15 (1) (2018)、p. 242、 10.1186/s12974-018-1282-6
- [127]
脱共役プロテイン 2 欠損は、in vitro および in vivo での高血糖による脳虚血および再灌流障害の増悪後の NLRP3 インフラマソーム活性化を増強するNeurochem Res、46 (6) (2021)、pp. 1359-1371、 10.1007/s11064-021-03270-9
- [128]
アディポネクチンペプチドは、AMPK/GSK-3βを調節することにより、脳虚血再灌流損傷後の酸化ストレスとNLRP3インフラマソーム活性化を軽減します経験値Neurol.、329 (2020)、記事 113302、 10.1016/j.expneurol.2020.113302
- [129]
腸内細菌はトリメチルアミン N-オキシド経路を介して脳卒中の重症度に影響を与える細胞宿主微生物、29 (7) (2021)、pp. 1199-1208、 10.1016/j.chom.2021.05.002Epub 2021 6 16. PMID: 34139173; PMCID: PMC8288076
- [130]
一酸化窒素供与体(硝酸塩)、 L-アルギニン、または急性脳卒中に対する一酸化窒素合成酵素阻害剤コクラン データベース システムRev.、4 (4) (2017)、記事 CD000398、 10.1002/14651858.CD000398.pub2PMID: 28429459; PMCID: PMC6478181。
- [131]
急性虚血性脳卒中における酸化ストレスを軽減し、炎症性微小環境を調節することにより神経保護を目的としたマクロファージ偽装二酸化マンガンナノ粒子上級科学。 (Weinh.)、8 (20) (2021)、記事 e2101526、 10.1002/advs.202101526Epub 2021 8 26. PMID: 34436822; PMCID: PMC8529435
- [132]
虚血性脳卒中における血小板と一酸化窒素生体内変化の役割: ベンチからベッドサイドまでの翻訳的レビューオキシド。 Med Cell Longev.、2020 (2020)、記事 2979260、 10.1155/2020/2979260PMID: 32908630; PMCID: PMC7474795。
- [133]
低酸素性または虚血性脳損傷における一酸化窒素合成酵素Rev. Neurosci.、26 (1) (2015)、pp. 105-117、 10.1515/revneuro-2014-0041PMID: 25720056
- [134]
脳虚血性低酸素症後の結果と回復における重要な因子としての一酸化窒素依存性経路生体分子、11 (8) (2021)、p. 1097、 10.3390/biom11081097PMID: 34439764; PMCID: PMC8392725。
- [135]
脳虚血再灌流のマウスモデルにおけるポリ(ADP-リボース)合成酵素の活性化におけるペルオキシ亜硝酸塩と神経の一酸化窒素合成酵素の役割神経科学。 Lett.、248 (1) (1998)、41-44 ページ
- [136]
ポリ(ADP-リボース)ポリメラーゼ遺伝子破壊によりマウスが脳虚血に対する耐性を獲得ナット。医学、3 (10) (1997)、1089-1095 ページ
- [137]
虚血性脳損傷はポリ(ADP-リボース)ポリメラーゼの活性化によって媒介されるJ.セレブ。血流。 Metab.、17 (11) (1997)、pp. 1143-1151
- [138]
ラットにおけるPARP-1誘発神経血管ユニット損傷の軽減による脳虚血後の脳の保護Brain Res、1624 (2015)、9-18 ページ
- [139]
脳、心臓、腎臓、精巣虚血/再灌流傷害における ROS 媒介 NLRP3 インフラマソーム活性化オキシド。 Med Cell Longev.、2016 (2016)、記事 2183026
- [140]
糖尿病を伴う虚血性脳卒中における潜在的な治療法としての NLRP3 インフラマソームJ. Neuroinflamm.、16 (1) (2019)、p. 121
- [141]
NLRP3欠損は実験的虚血性脳卒中における神経血管損傷を改善するJ.セレブ。血流。メタタブ、34 (4) (2014)、pp. 660-667
- [142]
Nrf2/ARE経路は、脳虚血再灌流後のBV2細胞におけるROS誘導性のNLRP3インフラマソーム活性化を阻害するインフラム。 Res、67 (1) (2018)、pp. 57-65
- [143]
ペルオキシ亜硝酸塩は筋萎縮性側索硬化症の SOD1(G93A) マウスモデルにおいて NLRP3 インフラマソームカスケードを活性化しますモル。 Neurobiol.、55 (3) (2018)、pp. 2350-2361
- [144]
酸化ストレスと炎症の関係:Toll 様受容体フリーラジカル。バイオル。医学、48 (9) (2010)、1121-1132 ページ
- [145]
トール様受容体 4 の早期遮断により、げっ歯類およびヒトの脳虚血モデルにおけるミクログリアの炎症誘発性表現型によって引き起こされる ROS と炎症が軽減されるBr. J. Pharm.、176 (15) (2019)、pp. 2764-2779
- [146]
トール様受容体 4 の新規調節因子である MicroRNA-1906 は、マウスの実験的脳卒中後の虚血性損傷を改善するJ. Neurosci.、37 (43) (2017)、pp. 10498-10515
- [147]
陽電子放射断層撮影による脳虚血後の細胞増殖と炎症に対するトール様受容体 4 の役割のイメージングJ.セレブ。血流。メタタブ、36 (4) (2016)、pp. 702-708
- [148]
JLX001 は、TLR2/4-NF-κB シグナル伝達経路の阻害を介して pMCAO ラットの炎症反応と酸化ストレスを調節しましたNeurochem Res、44 (8) (2019)、pp. 1924-1938
- [149]
虚血性脳卒中の病態生理学: 酸化ストレスの役割カー。薬局。 Des.、26 (34) (2020)、pp. 4246-4260、 10.2174/1381612826666200708133912
- [150]
虚血性脳卒中における神経炎症メカニズム: 心塞栓性脳卒中、背景、および治療法に焦点を当てるInt J.Mol.科学、21 (18) (2020)、p. 6454、 10.3390/ijms21186454
- [151]
虚血性脳卒中の治療標的としてのミトコンドリアフリーラジカル。バイオル。医学、146 (2020)、45-58 ページ、 10.1016/j.freeradbiomed.2019.11.005
- [152]
アポトーシス、シスプラチン耐性、壊死、虚血性細胞死、脳卒中および心筋梗塞における体積調節アニオンチャネル、VSORおよびMaxi-Clの役割カー。上。メンバー、83 (2019)、pp. 205-283、 10.1016/bs.ctm.2019.03.001
- [153]
脳梗塞・虚血性脳卒中における神経損傷:メカニズムから治療まで(総説)Int J.Mol. Med.、49 (2) (2022)、p. 15、 10.3892/ijmm.2021.5070
- [154]
神経細胞死生理。 Rev.、98 (2) (2018)、pp. 813-880、 10.1152/physrev.00011.2017
- [155]
脳卒中における細胞死のメカニズムと新しい分子および細胞治療の選択肢CurrNeuropharmacol、16 (9) (2018)、pp. 1396-1415、 10.2174/1570159X16666180302115544
- [156]
実験的局所脳虚血に対するアストラガロシド IV: 前臨床証拠と考えられるメカニズムオキシド。 Med Cell Longev.、2017 (2017)、記事 8424326、 10.1155/2017/8424326
- [157]
出血性脳卒中の病態生理学における P2X7 受容体関連プログラム細胞死CurrNeuropharmacol、16 (9) (2018)、pp. 1282-1295、 10.2174/1570159X16666180516094500
- [158]
脳卒中関連プログラム細胞死における炎症反応の役割CurrNeuropharmacol、16 (9) (2018)、pp. 1365-1374、 10.2174/1570159X16666180222155833
- [159]
CD137 リガンドと CD137 の相互作用は、虚血性脳卒中後のインフラマソーム関連脳損傷に必要です神経分子医学、22 (4) (2020)、pp. 474-483、 10.1007/s12017-020-08623-1
- [160]
脳卒中と免疫システム: 新しい戦略のレビューFolia Med (プロブディフ)、62 (3) (2020)、pp. 431-437、 10.3897/formed.62.e49451
- [161]
TREM2、ミクログリア、および神経変性疾患トレンド医学、23 (6) (2017)、512-533 ページ、 10.1016/j.molmed.2017.03.008
- [162]
神経炎症の発症におけるトール様受容体J. Neuroimmunol.、332 (2019)、16-30 ページ、 10.1016/j.jneuroim.2019.03.012
- [163]
虚血性脳卒中後の神経血管修復のための骨髄細胞上の潜在的な免疫療法標的Front Neurosci.、13 (2019)、p. 758、 10.3389/fnins.2019.00758
- [164]
NF-κB および MAPK シグナル伝達が虚血性脳卒中後のニューロンにおける NLRP インフラマソーム活性化を促進するという証拠モル。 Neurobiol.、55 (2) (2018)、pp. 1082-1096、 10.1007/s12035-017-0394-9
- [165]
カスパーゼ-1阻害は、脳虚血のマウスモデルにおけるパイロトーシスの標準的なインフラマソーム経路を標的とすることによりニューロン死を防止するCNS神経科学。 Ther.、26 (9) (2020)、pp. 925-939、 10.1111/cns.13384
- [166]
長いノンコーディングRNA MEG3はmiR-485/AIM2軸を標的とすることでピロトーシスを増加させ、脳虚血再灌流障害を促進する経験値Neurol.、325 (2020)、記事 113139、 10.1016/j.expneurol.2019.113139
- [167]
星状細胞の生理病理学: 細胞間ネットワーク、炎症、細胞死の交差点にある2015プログレ。 Neurobiol.、130 (2015)、pp. 86-120
- [168]
脳卒中における反応性アストログリオーシス:神経機能の回復に対するアストロサイトの寄与Neurochem Int、107 (2017)、pp. 88-103、 10.1016/j.neuint.2016.12.016
- [169]
乳児ニューロンセロイドリポフスチン症におけるアストロサイト活性化の低下の役割J. Neurosci.、31 (43) (2011)、pp. 15575-15585、 10.1523/JNEUROSCI.3579-11.2011
- [170]
IL-1β/IL-18プロセシングインフラマソームと他の炎症性サイトカイン間の相互調節カー。意見。免疫学、23 (5) (2011)、pp. 591-597、 10.1016/j.coi.2011.07.005
- [171]
虚血性脳卒中後の炎症: 白血球とグリア細胞の役割経験値Neurobiol.、25 (5) (2016)、pp. 241-251、 10.5607/en.2016.25.5.241
- [172]
NLRP2は虚血性脳卒中のマウスモデルで高度に発現されている生化学生物物理学。 Res Commun.、479 (4) (2016)、pp. 656-662、 10.1016/j.bbrc.2016.09.157
- [173]
ヒスピドリンは、NLRP3媒介ピロトーシスの抑制を通じて脳虚血再灌流障害に対して神経保護活性を示すLife Sci.、232 (2019)、記事 116599、 10.1016/j.lfs.2019.116599
- [174]
ラットの脳虚血/再灌流(I/R)損傷におけるNLRP6の効果J.Mol. Neurosci.、69 (3) (2019)、pp. 411-418、 10.1007/s12031-019-01370-4
- [175]
アストロサイトで発現する NLRP6 は、インフラマソームの活性化とピロトーシスの誘導を通じて、OGD/R 後のニューロン損傷を悪化させるInt Immunopharmacol.、80 (2020)、記事 106183、 10.1016/j.intimp.2019.106183
- [176]
虚血性脳卒中における神経炎症:MicroRNAを介したミクログリアの分極化に焦点を当てるフロントモールNeurosci.、13 (2021)、記事 612439、 10.3389/fnmol.2020.612439
- [177]
虚血性脳卒中における末梢単球とマクロファージの役割ニューロール。 Sci.、41 (12) (2020)、pp. 3589-3607、 10.1007/s10072-020-04777-9
- [178]
アストロサイトで発現する NLRP6 は、インフラマソームの活性化とピロトーシスの誘導を通じて、OGD/R 後のニューロン損傷を悪化させるInt Immunopharmacol.、80 (2020)、記事 106183、 10.1016/j.intimp.2019.106183
- [179]
NLRC4 インフラマソームが虚血性脳卒中後のアポトーシスおよびパイロトーシスによるミクログリア死を媒介するという証拠脳の行動。 Immun.、75 (2019)、34-47 ページ、 10.1016/j.bbi.2018.09.001
- [180]
ミクログリアのTREM-1受容体は実験的虚血性脳卒中においてSYKとの相互作用を介して神経炎症性損傷を媒介するCell Death Dis.、10 (8) (2019)、p. 555、 10.1038/s41419-019-1777-9
- [181]
二本鎖DNA感知cGASの阻害により虚血性脳卒中後の脳損傷が改善されるエンボモル。 Med、12 (4) (2020)、記事 e11002、 10.15252/emm.201911002
- [182]
脳虚血性損傷の過程における神経血管単位の役割薬局。レス、160 (2020)、記事 105103、 10.1016/j.phrs.2020.105103
- [183]
虚血性脳卒中における脳血管疾患と神経血管損傷円レス、120 (3) (2017)、pp. 449-471、 10.1161/CIRCRESAHA.116.308427
- [184]
虚血性脳卒中における血液脳関門損傷の神経炎症メカニズム午前。 J.Physiol. Cell Physiol.、316 (2) (2019)、pp. C135-C153、 10.1152/ajpcell.00136.2018
- [185]
脳卒中後の神経保護の潜在的なアプローチとしての NLRP3 インフラマソームの阻害科学。議員、8 (1) (2018)、p. 5971、 10.1038/s41598-018-24350-x
- [186]
新規 PGC-1α 活性化剤としてのメディオレシノールは、PPARα-GOT1 軸を介して虚血性脳卒中における内皮細胞のピロトーシスを防止します薬局。レス、169 (2021)、記事 105640、 10.1016/j.phrs.2021.105640
- [187]
オクルディンの分解により、in vitro で脳微小血管内皮細胞が再灌流損傷を受けやすくなるJ. Neurochem、156 (3) (2021)、pp. 352-366、 10.1111/jnc.15102
- [188]
カスパーゼ-1の阻害はパイロトーシスの活性化を抑制することで虚血に関連した血液脳関門の機能不全と統合性を改善するFront Cell Neurosci.、14 (2021)、記事 540669、 10.3389/fnsel.2020.540669
- [189]
カスパーゼ-1阻害は、M2ミクログリア/マクロファージを極性化し、NF-κB活性化を抑制することにより、実験的脳卒中における神経保護を媒介する生化学生物物理学。 Res Commun.、513 (2) (2019)、pp. 479-485、 10.1016/j.bbrc.2019.03.202
- [190]
脳卒中後の神経保護の潜在的なアプローチとしての NLRP3 インフラマソームの阻害科学。議員、8 (1) (2018)、p. 5971、 10.1038/s41598-018-24350-xPMID: 29654318; PMCID: PMC5899150。
- [191]
星状細胞由来細胞外小胞のMircoRNA-29aはTP53INP1およびNF-κB/NLRP3軸を介して脳虚血再灌流障害を抑制する細胞モル。 Neurobiol.、42 (5) (2022)、pp. 1487-1500、 10.1007/s10571-021-01040-3
- [192]
TREM-1 を標的とする LP17 は酸化ストレスとパイロトーシスを阻害することで脳虚血誘発性神経損傷を軽減する生化学生物物理学。 Res Commun.、529 (3) (2020)、pp. 554-561、 10.1016/j.bbrc.2020.05.056
- [193]
ヒスピドリンは、NLRP3媒介ピロトーシスの抑制を通じて脳虚血再灌流障害に対して神経保護活性を示すLife Sci.、232 (2019)、記事 116599、 10.1016/j.lfs.2019.116599
- [194]
低密度リポタンパク質受容体(LDLR)は、脳虚血/再灌流損傷後のNLRP3媒介神経ピロトーシスを制御するJ. Neuroinflamm.、17 (1) (2020)、p. 330、 10.1186/s12974-020-01988-x
- [195]
新規 PGC-1α 活性化剤としてのメディオレシノールは、PPARα-GOT1 軸を介して虚血性脳卒中における内皮細胞のピロトーシスを防止します薬理学。レス、169 (2021)、記事 105640、 10.1016/j.phrs.2021.105640
- [196]
CHRFAM7A の過剰発現は、NLRP3/カスパーゼ-1 経路によって媒介されるミクログリアのピロトーシスを阻害することで脳虚血再灌流障害を軽減する炎症、44 (3) (2021)、pp. 1023-1034、 10.1007/s10753-020-01398-4
- [197]
TP53 誘導性解糖およびアポトーシス調節因子は、低酸素/虚血誘導性ミクログリアのパイロトーシスおよび虚血性脳損傷を軽減します神経再生。 Res、16 (6) (2021)、pp. 1037-1043、 10.4103/1673-5374.300453
- [198]
骨髄間葉系幹細胞由来のエキソソームは、ミクログリアM1/M2表現型を調節することにより、脳虚血再灌流傷害誘発性の神経炎症およびパイロトーシスを軽減する経験値Neurol.、341 (2021)、記事 113700、 10.1016/j.expneurol.2021.113700
- [199]
低酸素状態で事前調整された嗅粘膜間葉系幹細胞は、HIF-1αを活性化することにより、脳虚血/再灌流誘発性のピロトーシスおよびミクログリア細胞のアポトーシス死を無効にするエイジング (ニューヨーク州アルバニー)、12 (11) (2020)、pp. 10931-10950、 10.18632/エイジング.103307
- [200]
ガストロジンは、抗酸化作用と抗炎症作用を改善し、アポトーシス経路を阻害することにより、マウスの脳虚血損傷を軽減します。Neurochem Res.、40 (4) (2015)、pp. 661-673、 10.1007/s11064-015-1513-5Epub 2015 1 月 13 日。PMID: 25582916
- [201]
ガストロジンは、抗酸化作用と抗炎症作用を改善し、アポトーシス経路を阻害することにより、マウスの脳虚血損傷を軽減します。Neurochem Res、40 (4) (2015)、pp. 661-673、 10.1007/s11064-015-1513-5
- [202]
ガストロジン前処理は脳虚血再灌流によって引き起こされるラットの脳損傷を軽減しますBrain Res、1712 (2019)、pp. 207-216、 10.1016/j.brainres.2019.02.006
- [203]
ガストロジンは、NLRP3/カスパーゼ-1経路を調節することにより、微小血管再灌流傷害誘発性ピロトーシスを改善しますJ.Physiol. Biochem、75 (4) (2019)、pp. 531-547、 10.1007/s13105-019-00702-7
- [204]
ガストロジンはlncRNA NEAT1/miR-22-3p軸を調節してピロトーシスを抑制し、脳虚血/再灌流障害を軽減するNeurochem Res、46 (7) (2021)、pp. 1747-1758、 10.1007/s11064-021-03285-2
- [205]
イー・シャオボさん。袁麗邦らNL RP3 インフラマソームシグナル伝達経路を介したグルコース酸素欠乏/再灌流損傷からのミクログリアに対するタンシノン IIA の保護効果J. 四川大学(医学編)、47 (5) (2016)、pp. 660-664
- [206]
慢性脳虚血ラットの認知機能に対するレオヌリンの影響顎。パット。医学、40 (9) (2018)、2045-2048 ページ
- [207]
関節リウマチ治療におけるシャクヤクの総グルコシドの薬理効果とメカニズム医学。彼女、34 (2) (2015)、199-201 ページ
- [208]
ペオニフロリンは酸に敏感なイオンチャネルを調節して鎮痛効果を発揮します医学。彼女、38 (11) (2019)、1403-1407 ページ
- [209]
急性脳梗塞ラットにおけるミクログリア活性化によって調節される神経細胞のピロトーシスに対するペオニフロリンの効果J.脳神経細胞。 Dis.、29 (08) (2021)、pp. 469-474
- [210]
NLRP3インフラマソームシグナル伝達経路を介したラットの脳虚血再灌流傷害後の細胞ピロトーシスに対するブチルフタリドの影響J.Integrate.顎。西。医学。心臓血管。脳血管。 Dis.、18 (06) (2020)、pp. 898-902
- [211]
NLRP3 インフラマソームの活性化を調節し、慢性脳虚血ラットの神経細胞のピロトーシスとアポトーシスを阻害します。J.安徽医学。大学、55 (09) (2020)、pp. 1321-1326、 10.19405/j.cnki.issn1000-1492.2020.09.002
- [212]
ラット脳組織における虚血再灌流中の細胞パイロトーシスに対するレスベラトロールの調節効果と、ミクログリアNLRP3インフラマソーム、カスパーゼ-1およびZO-1に対するその効果J.海南医学。コレクション、25 (17) (2019)、pp. 1291-1294、 10.13210/j.cnki.jhmu.20190716.002
- [213]
注射用サルビアノール酸は、in vivo および in vitro でミクログリアの M1/M2 表現型を切り替え、NLRP3 インフラマソーム/ピロトーシス軸を阻害することにより、脳虚血/再灌流損傷を軽減します。J. Ethnopharmacol.、270 (2021)、記事 113776、 10.1016/j.jep.2021.113776
- [214]
ヒスピドリンは、NLRP3媒介ピロトーシスの抑制を通じて脳虚血再灌流障害に対して神経保護活性を示すLife Sci.、232 (2019)、記事 116599、 10.1016/j.lfs.2019.116599
- [215]
Astragalus membranaceus 由来のアストラガロシド IV: 薬理効果に関する研究レビュー上級Pharmacol.、87 (2020)、pp. 89-112、 10.1016/bs.apha.2019.08.002
- [216]
心不全におけるアストラガロシド IV の最新の役割バイオメッド。薬学、126 (2020)、記事 110012、 10.1016/j.biopha.2020.110012
- [217]
がん治療におけるアストラガロシド IV の分子機構 (総説)Int J.Mol.医学、47 (3) (2021)、p. 13、 10.3892/ijmm.2021.4846
- [218]
心血管疾患の治療に効果的な薬薬物の研究DevelTher、14 (2020)、pp. 3731-3746、 10.2147/DDDT.S272355
- [219]
虚血性脳卒中および標的薬物療法における細胞死経路翻訳。脳卒中研究、11 (6) (2020)、pp. 1185-1202、 10.1007/s12975-020-00806-z
- [220]
体系的薬理学に基づいた HIF-VEGF 経路および脳虚血再灌流傷害の生物学的ネットワークに対する HedysarumMultijugumMaxim.-Chuanxiong Rhizoma 化合物の制御機構の探索Front Pharmacol.、12 (2021)、記事 601846、 10.3389/fphar.2021.601846
- [221]
定量的プロテオミクスによる脳梗塞に対する hedysarummultijugum maxim-chuanxiong 根茎化合物の神経保護効果の解明エヴィド。ベースの補完代替。医学、2022 (2022)、記事 5241902、 10.1155/2022/5241902
- [222]
hedysarummultijugummaxim.-Chuanxiong rhizoma 化合物の虚血性脳卒中に対する効果:ネットワークと実験薬理学に基づく研究オキシド。医学。 Cell Longev.、2020 (2020)、記事 6072380、 10.1155/2020/6072380
- [223]
脳虚血再潅流障害におけるマイトファジー-NLRP3 インフラマソーム経路の役割と、オタネニンジンサポニンとアストラガロシド IV の介入[D]湖南大学伝統。顎。医学。 (2019)、 10.27138/d.cnki.ghuzc.2019.000014
- [224]
低酸素性虚血性脳損傷を改善するためのMMP-9媒介NLRP3/カスパーゼ-1シグナル伝達経路を調節するアストラガロシドIVの分子機構[D]遼寧大学伝統。顎。医学。 (2021)、 10.27213/d.cnki.glnzc.2021.000042
- [225]
オタネニンジンサポニン:抗うつ作用または抗不安作用のメカニズムのレビューと植物化学および薬理学に関するネットワーク分析分子、23 (4) (2018)、p. 940、 10.3390/分子23040940
- [226]
オタネニンジンサポニンと神経系疾患におけるその応用:物語的レビューアン。翻訳。 Med、8 (22) (2020)、p. 1525年 10.21037/atm-20-6909
- [227]
オタネニンジンとオタネニンジンの化学成分が治療効果の違いを説明する薬理学。レス、161 (2020)、記事 105263、 10.1016/j.phrs.2020.105263
- [228]
ノトジンセンサポニンは全脳虚血後の海馬CA1領域における神経細胞のアポトーシスを抑制するおよびそのメカニズム顎。ハーブ。医学、47 (08) (2016)、pp. 1337-1344
- [229]
全脳虚血の成体ラットの側脳室の上衣領域における神経再生に対するオタネニンジンサポニンの効果顎。ハーブ。医学、47 (09) (2016)、pp. 1535-1540
- [230]
全脳虚血ラットにおける海馬大脳浮腫およびGFAP発現に対するオタネニンジンサポニンの効果顎。ハーブ。医学、48 (22) (2017)、pp. 4695-4700
- [231]
全脳虚血ラットの学習と記憶に対するオタネニンジンサポニンの効果とその作用機序顎。ハーブ。 Med.、50 (22) (2019)、pp. 5521-5526
- [232]
脳虚血再潅流障害におけるマイトファジー-NLRP3 インフラマソーム経路の役割と、オタネニンジンサポニンとアストラガロシド IV の介入[D]湖南大学伝統。顎。医学。 (2019)、 10.27138/d.cnki.ghuzc.2019.000014
- [233]
DRP1/NLRP3経路に基づく急性脳梗塞後のパイロトーシスに対するタオホンシウ煎じ薬の効果とメカニズム[D]北京大学伝統。顎。医学。 (2021)、 10.26973/d.cnki.gbjzu.2021.000498
- [234]
脳虚血再灌流ラットにおける細胞ピロトーシスに対するタオホンシウ煎じ薬の調節効果に関する研究 [D]安徽大学伝統。顎。医学。 (2020)、 10.26922/d.cnki.ganzc.2020.000120
- [235]
柿の葉フラボノイドの抗炎症および抗酸化神経保護効果 D-ガラクトース誘発老化マウスフダン J.: Med.編、42 (1) (2015)、7-12 ページ
- [236]
L-NAME誘発性高血圧ラットに対する柿の葉由来の総フラボノイドの降圧効果とメカニズム [D]広西医科大学、南寧市 (2008)
- [237]
心血管疾患と糖尿病の予防と治療における Ye とその製品の新たな進歩山西省地中海。 J.、37 (16) (2008)、740-741 ページ
- [238]
脳虚血に対する柿の葉有効成分と直心清錠の神経保護効果に関する研究[D]中山大学、広州 (2004)
- [239]
脳虚血再灌流障害モデルラットにおけるNaoxinqingの保護機構顎。組織工学Res.、24 (02) (2020)、pp. 215-222
- [240]
液体クロマトグラフィー四重極飛行時間型質量分析法を使用した、ラット尿中の芙蓉黄湯煎じ薬の代謝物の同定J.Pharm.バイオメッド。 Anal.、67–68 (2012)、pp. 114-122、 10.1016/j.jpba.2012.04.026
- [241]
Buyang Huanwu煎じ薬は、ラットのCRPとCD40を低下させることにより、気欠乏を伴う冠状動脈性心疾患とうっ血症候群を改善しますJ. Ethnopharmacol.、130 (1) (2010)、pp. 98-102、 10.1016/j.jep.2010.04.017
- [242]
hedysarum multijugum maxim.-Chuanxiong rhizoma 化合物の虚血性脳卒中に対する効果: ネットワークと実験薬理学に基づく研究オキシド。医学。 Cell Longev.、2020 (2020)、記事 6072380、 10.1155/2020/6072380
- [243]
HIF-VEGF経路および脳虚血再灌流傷害の生物学的ネットワークにおけるhedysarum multijugum maxim.-chuanxiong根茎化合物の制御機構を系統的薬理学に基づいて探索するFront Pharm.、12 (2021)、記事 601846、 10.3389/fphar.2021.601846
- [244]
ラットの脳虚血再灌流傷害後のパイロトーシスに対するBuyangHuanwu煎じ薬に含まれる配糖体の神経保護効果J. Ethnopharmacol.、242 (2019)、記事 112051、 10.1016/j.jep.2019.112051
- [245]
ラットの脳虚血再灌流傷害後のカスパーゼ-1およびIL-18タンパク質発現に対する龍芝煎じ薬の影響J.湖南大学伝統。顎。医学、41 (04) (2021)、pp. 518-522
- [246]
虚血性脳卒中回復期患者に対するYiqi Huoxue Tongluo煎じ薬の効果、および血行動態レベル、NO、VEGFおよびET-1レベルに対するその効果四川の伝統。顎。医学、37 (02) (2019)、pp. 142-144
- [247]
気を養い、血液を活性化するこの方法は、ピロトーシスを阻害することで急性虚血性脳卒中を患うラットの脳組織の炎症を軽減しますJ. 広州大学伝統。顎。 Med.、39 (01) (2022)、pp. 152-158、 10.13359/j.cnki.gzxbtcm.2022.01.026
- [248]
虚血性脳卒中に対する神納富源煎じ薬と星直開橋鍼治療の併用の臨床観察ワールド J. インテグレーションMed.、15 (01) (2020)、pp. 160-164+169、 10.13935/j.cnki.sjzx.200135
- [249]
MCAO ラットにおける Nogo-A および GAP-43 の発現に対する、hUC-MSC と組み合わせた神納富源煎じ薬の効果[D]湖南大学伝統。顎。医学。 (2021)、 10.27138/d.cnki.ghuzc.2021.000154
- [250]
細胞パイロトーシスのメカニズムに基づいた、グルコースと酸素が欠乏した PC12 細胞に対する神納富源煎じ薬の保護効果に関する研究 [D]湖南大学伝統。顎。医学。 (2019)、 10.27138/d.cnki.ghuzc.2019.000152
- [251]
敗血症誘発肺傷害のヘパリン治療の新しい経路の可能性: hMGB1-LPS 誘発カスパーゼ 11 活性化の遮断による肺内皮細胞ピロトーシスの阻害フロントセル感染。微生物、12 (2022)、記事 984835、 10.3389/fcimb.2022.984835PMID: 36189354; PMCID: PMC9519888。
- [252]
実験的心筋梗塞における柔らかいココナッツウォーターの抗酸化作用と抗血栓作用J. Food Biochem.、35 (5) (2011)、pp. 1501-1507
- [253]
若いココナッツジュース、アルツハイマー病に関連する一部の病状を大幅に軽減できる可能性のある治療薬:新たな発見Br. J. Nutr.、105 (5) (2011)、pp. 738-746、 10.1017/S0007114510004241Epub 2010 11 30。PMID: 21114897
- [254]
の薬力学 L-アルギニン変更。それで。 Health Med、20 (3) (2014)、pp. 48-54PMID: 24755570
- [255]
L-アルギニンと心血管系薬局。議員、57 (1) (2005)、14-22 ページPMID: 15849373
- [256]
抗血栓作用 L-高血圧ラットのアルギニンJ.Physiol.薬学、55 (3) (2004)、pp. 563-574PMID: 15381827
- [257]
アルギニンの静脈内投与は、NLRP3 インフラマソーム活性を下方制御し、多微生物性敗血症マウスの急性腎損傷を軽減する媒介。炎症、2020 (2020)、記事 3201635、 10.1155/2020/3201635PMID: 32454788; PMCID: PMC7238342。
引用者 (7)
-
カイコバトリチカトゥス抽出物は凝固因子Ⅻを活性化し、脳虚血/再灌流損傷を患ったラットの血管新生を促進します
2024年、民族薬理学ジャーナル -
脳虚血・再潅流後のミクログリアピロトーシスに対するジエドゥフオシュエ煎じ薬の神経保護効果—グリア血管単位の観点から
2024年、民族薬理学ジャーナル -
PI3K/Aktシグナル伝達を標的とした虚血性脳卒中における植物療法の進歩
2023年、植物療法研究